











-
酸性矿山废水(AMD)是含硫的矿物在氧气和水共同作用下,经风化、淋溶和微生物共同作用而形成[1].酸性矿山废水具有 低pH 值,高电导率,硫酸盐和重金属浓度高的特点[2],这对大多数生物来说都是难以生存的极端环境[3-5],然而这种极端环境胁迫下的独特生境却孕育着大量的微生物。大部分酸性矿山废水未经处理便直接排放到自然环境中,该环境下的微生物可以反过来影响外界水环境,微生物与水体的密切联系使得酸性废水水体生态修复成为可能.贵州省煤矿矿山规模偏小,分布面广,导致废水排放分散、持久,对山区水环境的污染相当严重[6].因此研究酸性矿山废水影响下环境中微生物群落结构特征及其与环境因子的关系具有重要意义.
微生物在水生生态系统中起着重要作用,其中浮游细菌扮演着多重角色(分解者、转化者、储存者和生产者)[7],与自然界中的碳、氮、磷、硫和微量元素铁等循环关系密切[8],在自然环境的自净过程中占有重要地位,是控制水生生态过程的关键因子,微生物食物网的关键组成部分[9].目前,越来越多的研究开始关注水体中的浮游细菌及其与环境因子之间的关系.研究者在国内外对不同纬度、不同大小的湖、库水体细菌群落结构组成开展了大量的研究工作[10-15],但主要集中在中性或碱性水库,对酸性水库细菌群落结构组成的研究鲜见报道. 目前在国际上,美国、西班牙、英国和法国等几个国家对AMD微生物多样性的研究得最广泛、最深入,其中最典型的两个研究样地为美国的IronMountain和西班牙的Tinto River. 然而,这些研究主要是基于湖库或者河流单一水生态系统的浮游细菌群落的调查,而对湖库及流域细菌群落结构和多样性的系统性调查研究还较为不足.浮游细菌与环境因子之间的关系较为复杂, 但在高通量测序技术的支撑下,能更加真实地揭示原位环境中微生物群落的复杂性和多样性[16],该方法摆脱了传统分离培养方法不能完整反映生态系统全部信息的束缚,逐渐成为各类环境中微生物群落结构和多样性研究的常用手段.
基于此,本研究选取贵州省兴仁市受矿区污染的青树子和马家屯水库及其入库流域作为研究区域,该区属亚热带温和湿润季风气候区,年均降水量约为1400 mm,年均气温为22.1 ℃.青树子水库(E:105°13'53.1'',N:25°23'12.7'',H:1371.8 m)及其入库流域,马家屯水库(E:105°08′35.3″,N:25°22′36.3″,H:1434 m)及其入库流域,它们的空间距离相对较近,地质背景条件相对一致(图1),特别是有含煤地层(P3l)的出露.该地区目前有一定规模的煤矿开采活动,矿山排出的酸性废水使得流域及水库水体产生不同程度的酸化.从上游到下游再从水库表层至底层进行系统采样,从水平空间再到垂向空间,调查不同污染梯度水体中细菌群落多样性的差异,使用MiSeq测序技术分析16S rDNA的丰度和多样性,并结合水质理化因子分析和主成分分析(PCA)对引起细菌群落结构和多样性变化的主要胁迫因子进行甄别,探讨了酸性废水影响下水库及入库流域水体中细菌群落结构特征及其影响因素,拓展了对酸性矿山废水污染环境中细菌群落多样性的了解.
-
水样采集于2019年9月,自流域上游至水库库底,共设置12个采样点,采样点分布见图1。青树子水库(QSZ水库)各垂直采样点为青树子水库表层(QSZS)、青树子水库中层(QSZM)和青树子水库底层(QSZB),其入库流域青树子流域(QSZ流域)上中下游采样点分别为QSZ-1、QSZ-2和QSZ-3,马家屯水库(MJT水库)各垂直采样点为马家屯水库表层(MJTS)、马家屯水库中层(MJTM)和马家屯水库底层(MJTB),其入库流域马家屯流域(MJT流域)上中下游采样点分别为MJT-1、MJT-2和MJT-3.
水库表层水样采自水面约5—15 cm处,中层水样采自水库深约7 m(青树子水库库深11.5 m,马家屯水库库深13.5 m)处,水库底部水样采至底泥上部。用有机玻璃容器采集 2 L 水样并混合均匀后,分装在预先灭菌的0.5 L 聚氯乙烯瓶中,分别用于理化指标和分子生物学分析测定.现场用滤膜(0.45 μm)过滤原水存储于50 mL离心管中,用于阴、阳离子和溶解性有机碳(DOC)测试,加入硝酸保存阳离子样品,加入磷酸保存DOC样品.再取1.5 L原水存储于已灭菌处理的聚乙烯瓶中,而后用0.22 μm(Milipore,美国)无菌微孔滤膜过滤水样,用以收集细菌,并将滤膜置于无菌离心管于干冰筒中低温(约–20 ℃)保存,运回实验室后进行后续处理.
-
水体温度(T)、pH、溶解氧(DO)和氧化还原电位(ORP)等参数在现场用多参数水质测定仪(Multi3630,德国WTW)测定.存储于500 mL棕色聚氯乙烯瓶中的水样总氮(TN)、氨氮(
${\rm{NH}}_4^+ $ -N)、总磷(TP)依据《水和废水监测分析方法》(第四版)[17]测定.存储于50 mL离心管中的水样K+、Ca2+、Na+、Mg2+、Mn2+、总铁(TFe)采用原子吸收测定(AFS-8510,美国赛默飞世尔),采用电感耦合等离子质谱仪测定(ICP-MS,美国赛默飞世尔)As3+、Cu2+、Pb2+、Cr2+,${\rm{SO}}_4^{2-}$ 使用离子色谱仪(ICS-1100,美国)测定,采用TOC分析仪(Analytikjena,德国)测定水体中DOC. -
冷冻保存的滤膜用无菌剪刀剪碎后采用E.Z.N.A Water DNA Kit基因组DNA试剂盒(Omega,USA)提取总DNA.测序委托生工生物工程(上海)股份有限公司进行,对微生物组总DNA提取,目标片段PCR扩增,扩增区域为标准细菌16S V3-V4,使用测试公司提供的通用引物341F(5'- CCTACGGGNGGCWGCAG–3')及805R(5'- GACTACHVGGGTATCTAATCC-3'),产物回收纯化、荧光定量及Illumina MiSeq高通量测序.
-
将Illumina MiSeq 测序得到的下机数据(Raw Data)进行处理得到Tags序列,并与数据库(Silva:
http://www.arb-silva.de/ )进行对比分析,检测并去除其中的嵌合体序列,得到最终的有效数据(Effective Tags).本文以 OTU 为单位,利用 Mothur(v1.30.1)软件进行群落多样性指标的统计分析;采用QIIME(v1.8.0)对生物信息学进行分析,根据序列的相似度,将序列归为多个OTU(operational taxonomic unit);为了得到每个 OTU 对应的物种分类信息,采用RDP classifier 贝叶斯算法分类学分析在 97%相似水平的 OTU代表序列,并统计各个分类水平每个样品的群落组成;采用主成分分析 (Principal component analysis,PCA)方法,运用 CANOCO(Version 5.0)软件解析浮游细菌群落与水质之间的关系,采用蒙特卡洛置换检验(Monte Carlopermutation text)统计环境因子与细菌群落的相关性[18-19]. -
水样的基本理化性质见表1.不同空间位点水体理化特征呈现不同变化趋势.受AMD影响的水库及入库流域水体中,pH在上游至下游整体呈增加趋势,水库表层至底层整体为下降趋势.其中QSZ流域水体pH值为3.51—4.02,QSZ水库水体pH值为4.60—4.78,水质为酸性.MJT流域水体pH为3.82—6.56, MJT水库水体pH值为4.24—6.13,水质酸性较弱.水温T在上游至下游逐渐增高,水库表层至底层逐渐降低,QSZ流域水体温度从上游20.4℃增加到下游22.3 ℃,QSZ水库水体温度从表层23.8 ℃降低至底层23.1 ℃,而MJT流域水体温度从上游21.3 ℃升高到下游24.8 ℃,MJT水库水体温度从表层23 ℃降低到底层21.8 ℃.EC、Eh、DO上游至下游逐渐下降,水库表层至底层EC、Eh变化趋势不明显,DO逐渐下降.
TN、TP、
${\rm{NH}}_4^+ $ -N、DOC营养物质在流域中变化趋势不明显,从水库表层至底层浓度逐渐增加,且在水库水体中的浓度整体低于入库流域水体,可能是因为河流是一个动态的水体,其水的流动性较湖库要高,水体的营养盐分布更为均质.其中,在QSZ-1和MJT-2处各营养物质浓度值均较大,TN、TP、${\rm{NH}}_4^+ $ -N、DOC营养物质分别为QSZ-1(1.80、4.88、0.98、2.48 mg.L−1),MJT-2(2.90、0.08、1.06、1.39 mg.L−1),且QSZ-2位点处TP达到最大值为5.86 mg.L−1.可能是因为水体中有大量生活垃圾倾倒和生活污水排放,从而导致了局部污染的急剧增加.整个沿程变化水体中各金属离子(Mg2+、Mn2+、As3+、K+、Na+、Cr2+、Cu2+、Pb2+)浓度均较低,流域水体中最大值低于250 mg.L−1,其中Mg2+、Mn2+、As3+上游至下游浓度逐渐降低,水库水体中各金属离子浓度无明显变化趋势,且各金属离子含量均低于100 mg·L−1.SO42–离子浓度在流域水体中的浓度要高于水库水体中,在QSZ-1和MJT-1处达最大值,分别为1065.65 mg.L−1和1570.19 mg.L−1. -
QSZ区域和MJT区域各选择6个代表性样点进行细菌群落结构多样性分析.表2为各水样有效序列数目、操作分类单位(OTUs)数目、多样性指数(Shannon、ACE、Chao1、Simpson)以及覆盖度指数(Coverage).QSZ水库及入库流域水样的高通量测序共得到518807条高质量序列,MJT水库及入库流域水样共获得498409条高质量序列.所有水样的覆盖度均在96%以上,说明测序深度可以较好的反映水体中浮游细菌的多样性和群落特征.以97%相似度划分,QSZ区域共得到23255个OTUs,每个水样平均OTUs数目为3876,MJT区域共得到22500个OTUs,单个水样平均OTUs数目为3750.可以看出两个区域的细菌群落多样性差异不明显.其中,QSZ区域OTU数目排在前两位的样品是QSZ-3和QSZB,最少的为QSZS,MJT区域中MJT-2是OTU数目最多的样品,最少的为MJTS.
Shannon指数和Simpson指数反映细菌多样性,Shannon指数值越大,群落多样性越高,Simpson指数所表现的多样性与Shannon指数相反.ACE指数和Chao1指数可估计细菌丰富度,其值越大,细菌丰富度越高.QSZ-3、QSZB和MJT-2的Shannon指数、ACE指数、Chao1指数值均较大,表明青树子流域下游、青树子水库底部以及马家屯水库中游采样点处的细菌丰富度与多样性均处于较高水平,而QSZS和MJTS的Shannon指数、ACE指数、Chao1指数值均偏小,表明青树子水库表层与马家屯水库表层采样点处的细菌丰富度与多样性水平均较低.水库水样OTUs和Shannon指数呈现出QSZS<QSZM<QSZB、MJTS<MJTM<MJTB的规律(图2)。
从水库表层到底层该段生态系统所承受的环境压力逐渐减小.而从流域上游到下游由于人类活动影响较为明显导致该段生态系统所承受的环境压力大小没有明显变化趋势。因此,从上游至水库底部,除在水库底部外,细菌群落多样性在流域水样中要高于水库水样,在流域中变化趋势不明显,但水库水样从表层至底层细菌多样性不断增加.表层光照时间充足,有利于浮游植物的生长,叶绿素含量高,使其对水体营养物质消耗,随着深度的增加,光照不足造成 BOD5 和叶绿素 a含量下降[20],底层水体营养物质更加丰富,为底层微生物提供了适宜的生存条件,从而浮游细菌多样性的垂直分布底层大于表层,造成微生物群落的空间差异性.Humayoun等[21]通过克隆文库法, 同样发现湖泊中好氧层和厌氧层间细菌多样性呈现显著差异, 细菌多样性在表层水体中较低.这可能与有机质沉降和水—底界面物质交换有关[22],这种作用的结果使得底层水体有机碳(POC)含量增高,从而使得底层细菌多样性高于表层.本研究中随着水深的增加,氮磷等营养物质含量和细菌多样性也在增大,且显示水库表层细菌香农指数最低,这与Humayoun等的研究结果相似.
-
QSZ区域所有水样浮游细菌共检测到37个门,MJT区域水样共检测到35个门,两个区域水体中相对丰度最高的4个门为变形菌门(Proteobacteria)、拟杆菌门(Bacteroidetes,QSZ:1.47%—12.96%,MJT:0.21%—11.81%)(下同)、厚壁菌门(Firmicutes,0.15%—14.48%,0.28%—10.57%)和放线菌门(Actinobacteria,0.06%—4.64%,0.89%—3.63%)(图3a),Proteobacteria中相对丰度占比较高的是α-变形菌纲(Alphaproteobacteria,2.70%—61.99%,2.97—57.15%)、β-变形菌纲(Betaproteobacteria,2.14%—63.53%,1.29%—42.67%)和γ-变形菌纲(Gammaproteobacteria,9.39%—63.09%,6.41%—45.32%).其中,Alphaproteobacteria相对丰度在两个区域呈现相同的变化规律,从上游至下游依次升高,且水库中层相对丰度最高,QSZM 为61.99%,MJTM为 57.15%,底层最低,QSZB为41.60%,MJTB为40.97%.Betaproteobacteria相对丰度在QSZ流域上游<中游<下游,而在MJT流域为上游>中游>下游,水库水样底层相对丰度高于表层和中层,QSZB为12.02%,MJTB为16.68%.Gammaproteobacteria相对丰度在QSZ流域中游达最高值,为63.09%,在MJT流域下游达最高值,为44.44%,水库表层至底层相对丰度依次降低.Bacteroidetes、Firmicutes在QSZ-1达QSZ区域最高值,分别为12.96%和14.48%,在MJT-2达MJT区域最高值,分别为11.81%和10.57%,水库表层至底层,相对丰度依次增加. Actinobacteria相对丰度在上游至下游依次降低,水库表层至底层依次增加.
研究结果发现,水体中的浮游细菌在门水平主要由Proteobacteria(Alphaproteobacteria、Betaproteobacteria和Gammaproteobacteria)、Bacteroidetes、Firmicutes、Actinobacteria组成,这与大量研究地表水细菌群落结构的文献相似[23-26].Proteobacteria通常是水环境中主要的细菌类群,包含了光营养性、化能自养型和化能异养型细菌,这类细菌在水环境中普遍存在[27-28],也是本研究中的优势门.不同的变形菌亚类在不同空间位置呈现不同的变化趋势.Alphaproteobacteria相对丰度从上游至下游依次升高.Betaproteobacteria广泛分布于淡水湖泊中,是迄今为止被研究最多的细菌种群之一[29],其在淡水湖中的大量存在与它们对生物可利用性元素含量变化的快速响应能力有关[30].Gammaproteobacteria作为Proteobacteria占比较大的组分之一,对有机化合物的降解起着重要的作用[31].Gammaproteobacteria在青树子流域中游和马家屯流域下游的相对丰度明显高于其他样品.有研究表明,Gammaproteobacteria通常存在于农业活动或者有机污染的地区[32].可能这两个位点较高的无机营养盐含量使得其相对丰度达到了最大值.Bacteroidetes和Firmicutes相对丰度在水库水体中随着深度的增加而升高. Actinobacteria相对丰度呈现出上游>中游>下游,底层>中层>表层的变化趋势,该类菌大部分属于异养型细菌,能分解多种有机物[33],本研究中氮磷含量在入库流域和水库底部水体中较高,可导致其水体中的Actinobacteria繁殖量增加[34],这可能是该菌类相对丰度在流域上游和水库底部较高的原因.
在属水平上,QSZ区域共检测出525个类群,相对丰度大于1%的有19个.MJT区域共检测出592个类群,相对丰度大于1%的有34个(图3b).QSZ区域排名前五的是红细菌属(Rhodobacter,0.01%—57.52%)、不动杆菌属(Acinetobacter,3.89%—57.84%)、盖氏铁柄杆菌属(Gallionella,0—38.45%)、假单胞菌属(Pseudomonas,0.28%—14.31%)和铁细菌属(Ferrovum,0.68%—5.04%).MJT区域排名前五的除Rhodobacter(0.22%—37.15%)、Acinetobacter,(2.13%—31.29%)、Gallionella(0—16.08%)和Ferrovum(0.01%—13.21%)外,还有甲基孢囊菌属(Methylocystis,0.02%—31.86%).QSZ区域上游水体细菌相对丰度较高(大于5%)的类群是Acinetobacter(21.69%)、Pseudomonas(12.29%)和Bacteroides(5.02%),中游水体相对丰度较高的是Acinetobacter(57.84%)和Gallionella(11.91%),下游水体相对丰度较高的是Gallionella(38.45%),水库表层水体中丰度较高的是Rhodobacter(55.96%)、Acinetobacter(19.75%)和Ferrovum(5.04%),中层水体中丰度较高的是Rhodobacter(57.52%)、Acinetobacter(16.49%)和Pseudomonas(14.31%),底层水体中丰度较高的是Rhodobacter(19.77%).而MJT区域上游水体细菌相对丰度较高(大于5%)的类群有Acinetobacter(17.09%)、Gallionella(16.08%)和Ferrovum(13.21%),中游水体中相对丰度较高的类群有噬氢菌属(Hydrogenophaga,5.94%)、新鞘脂菌属(Novosphingobium,5.34%),下游水体中相对丰度较高的类群有Rhodobacter(12.73%)、Acinetobacter(31.29%)、Methylocystis(31.86%)、军团菌(Legionella,7.62%),水库表层水体中相对丰度较高的类群有Rhodobacter(23.59%)、Acinetobacter(27.95%)、Methylocystis(21.33%)、Legionella(7.05%),中层水体中相对丰度较高的类群有Rhodobacter(37.15%)、Acinetobacter(22.65%)、Methylocystis(13.58%),底层水体中相对丰度较高的类群有Rhodobacter(28.57%).因此,受AMD影响的水库水体及其入库流域水体中,Acinetobacter自流域上游到水库底层均占较大比例,上游水体优势菌属有Pseudomonas、Ferrovum、Bacteroides ,中游水体优势菌属有Hydrogenophaga、Gallionella、Novosphingobium,而Rhodobacter是下游及水库水体中的优势菌属.
在属水平,优势类群主要有Rhodobacter、Acinetobacter、Gallionella、Pseudomonas、Ferrovum.Acinetobacter在受AMD影响的上游至水库底层水样中均占较大比例,Pseudomonas、Ferrovum和Bacteroides是上游水样中的优势菌属,Hydrogenophaga、Gallionella、Novo sphingobium是中游占比较大的菌属,Rhodobacter是水库下游、水库水样中的优势细菌.Rhodobacter能够通过光合作用进行生长代谢,CO2 和氮的固定,在水生生态系统的碳、氮循环中发挥重要作用[35-36].水库属于封闭型区域,外源营养盐供给较少,水体营养盐成为浮游细菌生长的限制因子,细菌的自养过程逐步扮演相对重要的角色,因而光合细菌相对丰度较入库流域水体增高.Bacteroides类群中的微生物广泛分布于厌氧环境中,并具有降解一系列复杂有机高分子的能力,包括碳水化合物及蛋白质[37].Wang 等[38]的研究表明城市段河流中Bacteroidetes 丰度较高的原因可能与居住区未经处理的排放废水中含有硝酸盐、氨、粪便等污染物有关.本研究中Bacteroidetes在青树子上游和马家屯中游处相对丰度达最大值,与该处附近有居民生活污水的排放有关,且水库底层丰度高于表层和中层,随着深度的增加,水库底部不断形成的拥有丰富有机物的厌氧环境使得Bacteroidetes适宜生存,相对丰度增加.Pseudomonas作为受AMD影响流域中游处的优势菌属之一,还是一种反硝化聚磷菌,在磷的转化和循环过程中也有重要作用.美国威斯康星大学 McLellan等[39]研究组通过比较分析污水细菌群落的宏基因组数据研究发现,Acinetobacter是污水细菌群落的核心类群,罗宁等[40]研究发现该菌属没有反硝化聚磷作用,但能在好氧条件下吸磷,而在青树子流域上游和中游水体总磷含量分别高达4.88 mg.L−1和5.86 mg.L−1,远高于地表水Ⅴ类水标准(0.4 mg.L−1),如此高的磷含量为Pseudomonas和Acinetobacter在该位点的代谢作用提供了较好的反应条件.
-
将细菌群落的Shannon指数、Simpson 指数和Chao1指数与水环境因子进行PCA分析(图4a),并通过蒙特卡洛显著性检验(Monte Carlopermutation text)进行统计,用箭头表示细菌群落与环境指标,箭头连线的长度表示细菌群落相对丰度与该环境指标相关系数的大小.结果表明,上中下游水体采样点主要集中在排序轴右侧,浮游细菌物种多样性主要与NH4+-N(F=8.3,P=0.032)、T(F=7.0,P=0.02)显著相关,均在P<0.05水平,解释度分别为45.3%、41.2%.因此,
${\rm{NH}}_4^+ $ -N是显著影响流域水体细菌群落的环境因子,而T对水库水体细菌群落显著影响.对属水平优势细菌与水环境因子进行PCA分析(图4b),由图可知,上、中、下游水体采样点主要集中在排序轴左侧,蒙特卡洛显著性检验表明,${\rm{NH}}_4^+ $ -N(F=6.4,P=0.002)、${\rm{SO}}_4^{2-} $ (F=3.6,P=0.026)、TP(F=2.9,P=0.03)是显著影响上下游水体中浮游细菌群落的环境因子,解释度分别为39.2%、33.4%、26.4%,T(F=2.6,P=0.048)是显著影响水库水体细菌群落的环境因子,解释度为20.5%,其中${\rm{NH}}_4^+ $ -N在P<0.01水平上显著影响,${\rm{SO}}_4^{2-} $ 、TP、T在P<0.05水平上显著影响.综上可知,受AMD影响的入库流域水体中${\rm{NH}}_4^+ $ -N、TP和${\rm{SO}}_4^{2-} $ 是驱动细菌群落空间变异的主要因子,受AMD影响水库水体中水温T是主要的驱动因素.此外,Mg2+、As3+对入库流域水体细菌群落也存在一定影响,但未达统计学显著水平.越来越多的研究表明, 温度是影响湖泊细菌群落结构和组成的关键因子之一[41-42],而本研究结果也表明水温T是受AMD影响水库水体中浮游细菌群落结构的重要调节因素.该结论与 Humayoun等研究结果相似,不同深度的湖泊微生物多样性,下层水样比上层水样高,这可能是由于库底缺氧、低温所造成的[43].李玉华等[44]在研究松花湖水体垂向空间细菌的多样性时也发现,从表层至底层随着水温T的降低,细菌群落多样性随之升高.由于本研究中的AMD环境与其余类似生态系统相比,上游有大量生活污水和垃圾的倾排,环境相对温和,致使入库流域的水体中细菌主要受氮磷的影响程度较大,其水体中的有效磷足以维持光自养生物的生长.有研究表明无机营养物可能是微生物生长的限制因子,其中尤其以磷含量对水体中微生物数量的影响最为明显[45],这种限制在AMD水体中可能会加剧,因为磷通常会与铁和铝羟化物在AMD环境中共沉淀[46].由于在低pH条件下抑制硝化作用,以氨形式存在的氮在AMD水体中也较充足[47].因此,高磷和高氮可能是主导该研究受AMD影响较为严重的入库流域水体中微生物群落变化的主要原因.PCA分析表明
${\rm{SO}}_4^{2-} $ 浓度对流域水体中细菌群落结构造成了显著影响,在本文水体理化性质分析中可知在流域水体中${\rm{SO}}_4^{2-} $ 浓度高于水库水体,因此,${\rm{SO}}_4^{2-} $ 同样对AMD影响的河流浮游细菌起到了不可忽略的作用,Zhang等[48]在研究博斯腾湖细菌群落时空变化时发现,高浓度的${\rm{SO}}_4^{2-} $ 提供了良好的营养条件,从而促进细菌的生长.此外,Mg2+、AS3+也与入库流域水体中的细菌群落产生了影响,但影响程度小于${\rm{SO}}_4^{2-} $ ,AMD生态系统中的微生物能够将重金属吸收到细胞内或吸收到细胞外多糖上[49],从而降低了重金属的溶解度,有利于其他原生微生物的生长.因此,浮游细菌群落特征不是由单一的某种环境因子决定的,而是由不同环境因子综合调控. -
研究结果不但帮助人们系统地认识了AMD环境中细菌多样性,还有利于了解细菌在AMD环境中的适应机制和生存状态,以期为酸性矿山废水区域的生态恢复提供理论基础.主要结论如下:
(1)受AMD影响下,入库流域水体中的细菌群落多样性无明显变化趋势,水库水体中细菌群落多样性由表层至底层逐渐增多.
(2)在门水平,受AMD影响水库及其入库流域水体中主要的细菌门类有Proteobacteria(Alphaproteobacteria纲、Betaproteobacteria纲和Gammaproteobacteria纲)、Bacteroidetes、Firmicutes、Actinobacteria.Alphaproteobacteria相对丰度从上游至下游依次升高.Betaproteobacteria在水库底层水体中相对丰度达最大值,而Gammaproteobacteria相对丰度随着水库深度的增加而降低.Bacteroidetes、Firmicutes在水库水体中随着深度的增加相对丰度升高.Actinobacteria相对丰度上游至下游逐渐降低,水库水体中表层至底层逐渐增高.在属水平,受AMD影响水库及其入库流域水体中优势类群主要有Rhodobacter、Acinetobacter、Gallionella、Pseudomonas、Ferrovum.Acinetobacter在上游至水库底层水样中均占较大比例,Pseudomonas、Ferrovum、Bacteroides是上游水样中的优势菌属,Hydrogenophaga、Gallionella、Novosphingobium在中游占比较大,Rhodobacter是下游水样及水库水样中的优势细菌.
(3)
${\rm{NH}}_4^+$ -N、TP和${\rm{SO}}_4^{2-} $ 是主导受AMD影响下入库流域水体中细菌群落结构变化的环境因子,水温T对受AMD影响水库水体中细菌群落造成了显著影响,此外,Mg2+、As3+对入库流域水体细菌群落也存在一定影响.
酸性矿山排水影响下水库及其入库流域水体细菌多样性与环境因子的关系
Relationship between bacterial diversity and environmental factors in reservoirs and their inflow basins under the influence of acid mine drainage
-
摘要: 为了解酸性矿山废水排水处沿流域至水库底层水体细菌群落结构的沿程变化,探索其环境影响因素.采用高通量测序技术并结合主成分分析(principal component analysis,PCA)方法,对2019年9月采集受酸性矿山废水(acid mine drainage,AMD)影响的12个水样进行研究.结果表明,入库流域水样中细菌群落多样性变化趋势不明显,从表层至底层细菌多样性不断升高.在门水平上受AMD影响的细菌主要由变形菌门(Proteobacteria)、拟杆菌门(Bacteroidetes,0.21%—12.96%)、厚壁菌门(Firmicutes,0.15%—14.48%)和放线菌门(Actinobacteria,0.06%—4.64%)构成。其中,变形菌门中的α-变形菌纲(Alphaproteobacteria,2.42%—61.99%)、β-变形菌纲(Betaproteobacteria,1.29%—63.53%)和γ-变形菌纲(Gammaproteobacteria,6.41%—63.09%)为优势菌类.从不同空间位置浮游细菌群落组成来看,不动杆菌属(Acinetobacter)在受AMD影响流域上游至水库底层水样中均占较大比例,为2.13%—57.84%,假单胞菌属(Pseudomonas,3.86%—12.29%)、铁细菌属(Ferrovum,0.97%—13.21%)和拟杆菌属(Bacteroides,0.01%—5.02%)是上游水样中的优势菌属,噬氢菌属(Hydrogenophaga,0—5.94%)、盖氏铁柄杆菌属(Gallionella,1.45%—11.91%)和新鞘氨醇杆菌(Novosphingobium,0—5.34%)是中游占比较大的菌属,红细菌属(Rhodobacter,12.73%—57.52%)是下游水样和水库水样中的优势细菌.造成AMD影响下不同位点处水体浮游细菌群落组成差异的环境因子各不相同,入库流域水体细菌群落主要受
${\rm{NH}}_4^+ $ -N、总磷(TP)和${\rm{SO}}_4^{2-}$ 影响,Mg2+和As3+也与入库流域水体中细菌多样性存在一定相关关系,水库水体中细菌群落主要受水温(T)影响.研究结果将有助于为酸性矿山废水微生物资源及生态环境的研究提供理论依据.Abstract: To understand the changes in bacterial community structure along the watershed to the bottom of the reservoir at the acid mine drainage and to explore the environmental impact factors. Twelve water samples affected by acid mine drainage (AMD) were collected in September 2019 using high-throughput sequencing technology and principal component analysis (PCA). The results showed that the trend of bacterial community diversity in the water samples from the inlet basin was not obvious, and the bacterial diversity increased from the surface to the bottom layer. The bacteria affected by AMD at the phylum level were mainly Proteobacteria, Bacteroidetes (0.21%—12.96%), Firmicutes (0.15%—14.48%), and Actinobacteria( 0.06%—4.64%). Of these, the phylum Alphaproteobacteria (2.42%—61.99%), the phylum Betaproteobacteria (1.29%—63.53%) and the phylum Gammaproteobacteria (6.41%—63.09%) were the dominant bacterial groups. In terms of the composition of the planktonic bacterial community at different spatial locations, Acinetobacter(2.13%—57.84%), Pseudomonas(3.86%—12.29%), Ferrobacter(0.97%—13.21%), and Bacteroides (0.01%—5.02%) were the most abundant species in the water samples from the upstream to the bottom of the reservoirs affected by AMD. Pseudomonas (3.86%—12.29%), Ferrovum (0.97%—13.21%) and Bacteroides (0.01%—5.02%) were the dominant genera in the upstream water samples, Hydrogenophaga (0—5.94%), Gallionella (1.45%—11.91%) and Novosphingobium(0—5.34%) were the dominant genera in the midstream, and Rhodobacter(12.73%—57.52%) were the dominant bacteria in the downstream water samples and in the reservoir water samples. The environmental factors leading to the difference in the composition of the planktonic bacterial community at different sites under the influence of AMD are different,${\rm{NH}}_4^+$ -N, total phosphorus (TP) and${\rm{SO}}_4^{2-} $ were the main factors affecting the bacterial community in the inflow basin. Mg2+ and As3+ also had a certain correlation with the bacterial diversity in the inflow basin, and bacterial community in reservoir water is mainly affected by water temperature (T). The results of the study will help to provide a theoretical basis for the study of the microbial resources and ecological environment of acid mine wastewater.-
Key words:
- acid mine drainage /
- bacterial diversity /
- environmental factors /
- watershed /
- reservoir /
- high-throughput sequencing
-

-
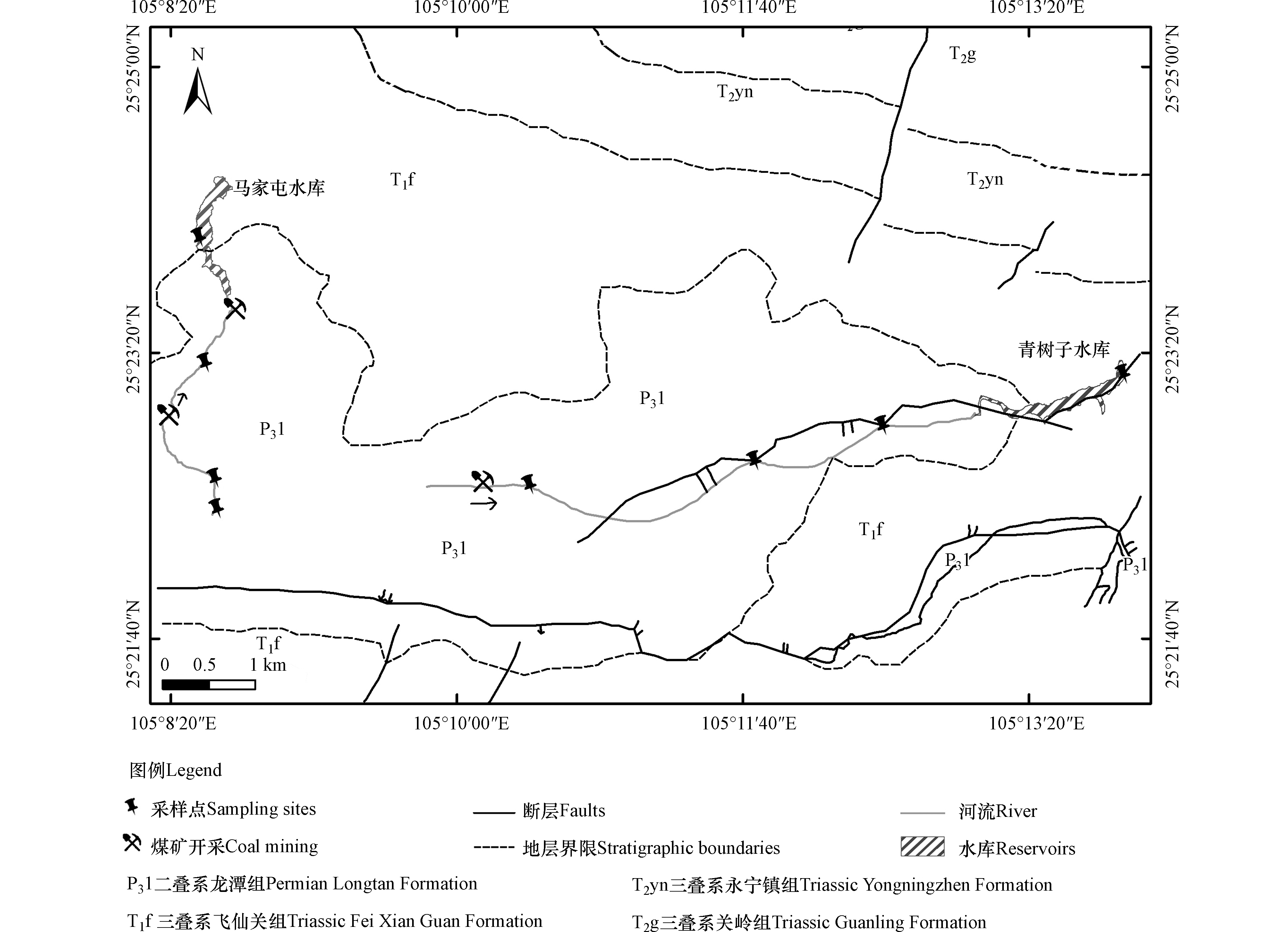
图 1 研究区地质环境及采样位置分布图
Figure 1. Map of geological environment and sampling sites in the study area
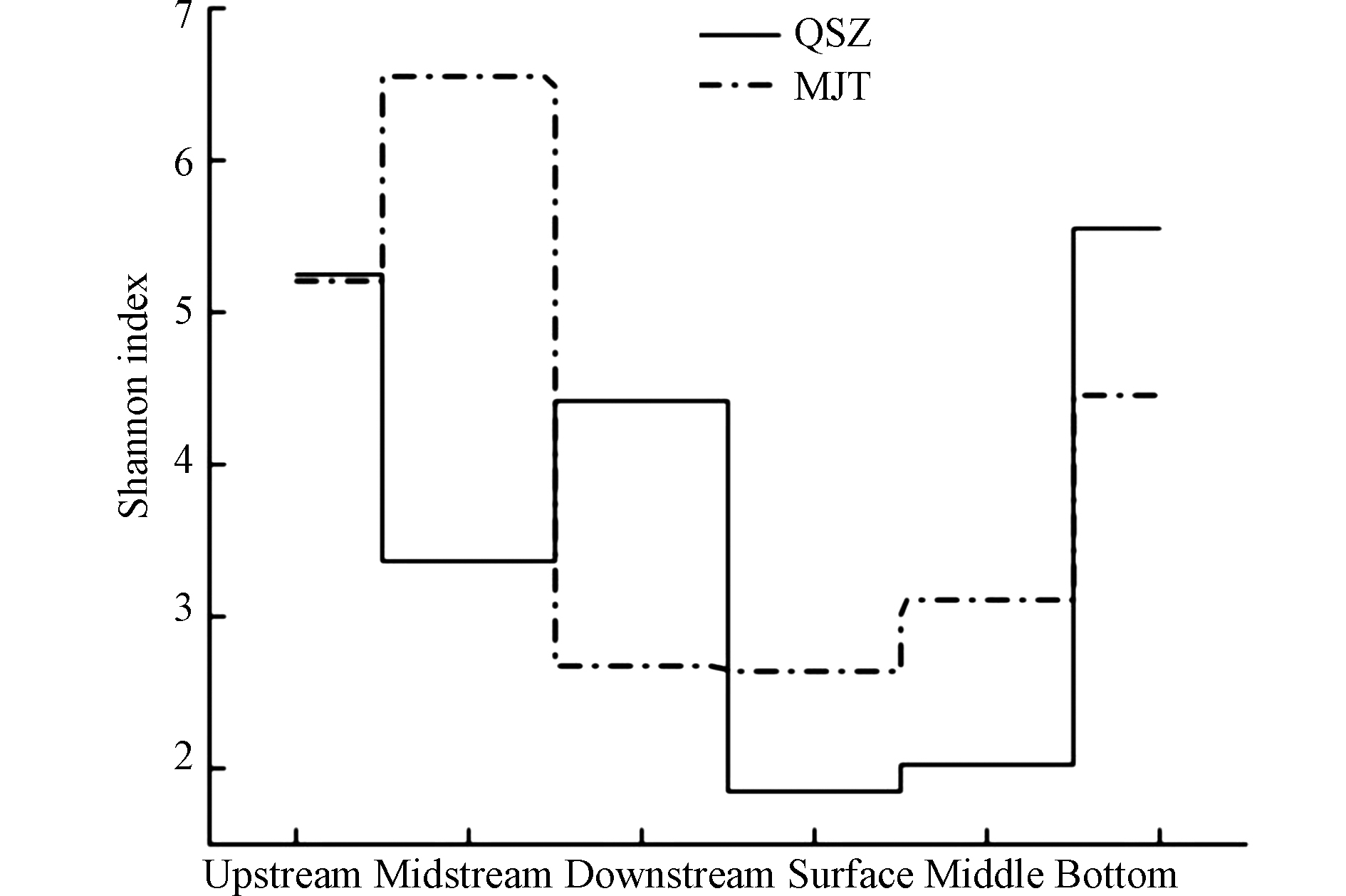
图 2 水体浮游细菌群落香农指数变化
Figure 2. Shannon index changes of phytoplankton bacterial community in water
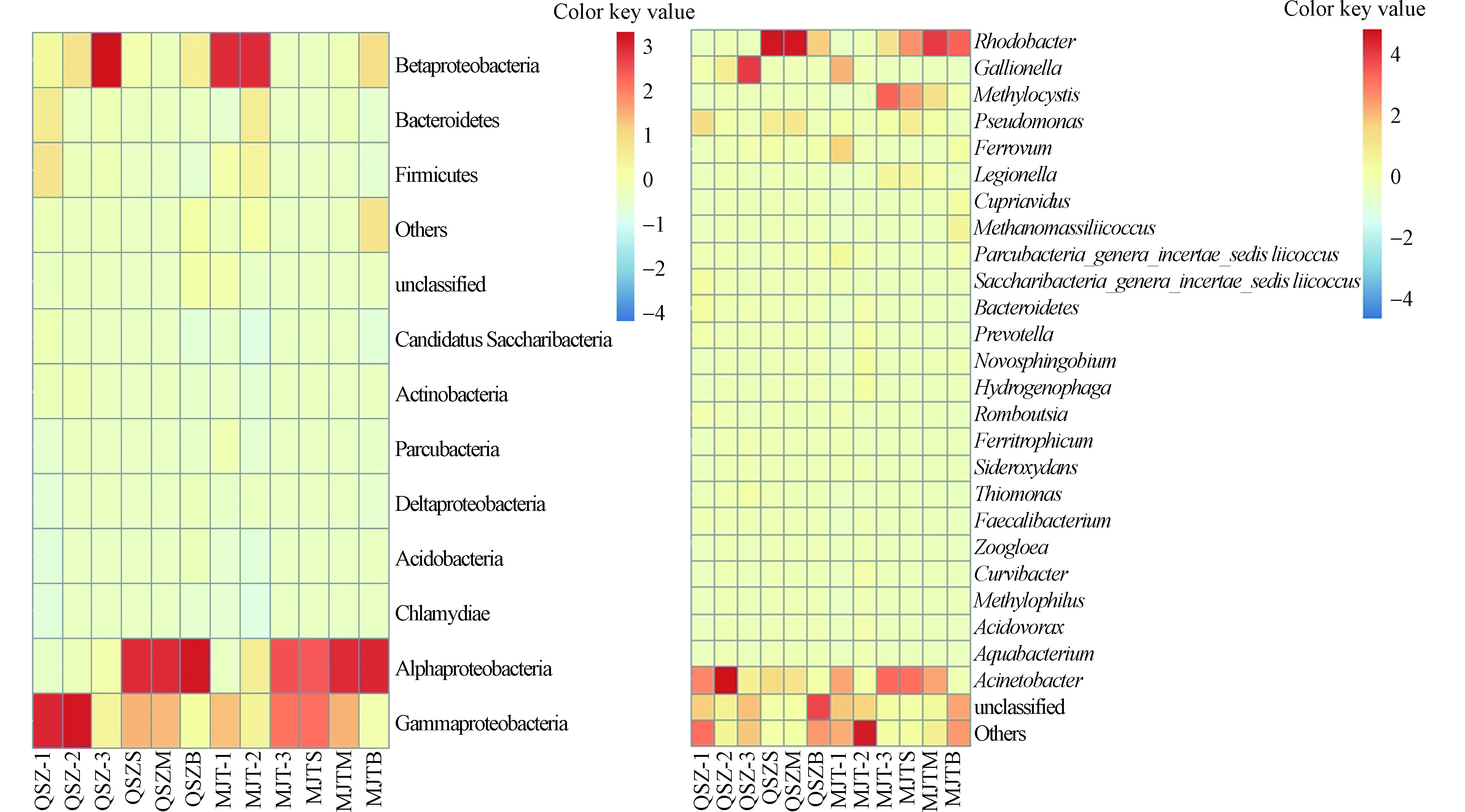
图 3 不同样品中细菌门水平(a)与属水平(b)上细菌群落结构与分布
Figure 3. Structure and distribution of bacterial community at both phylum level (a) and genus level (b) in different samples
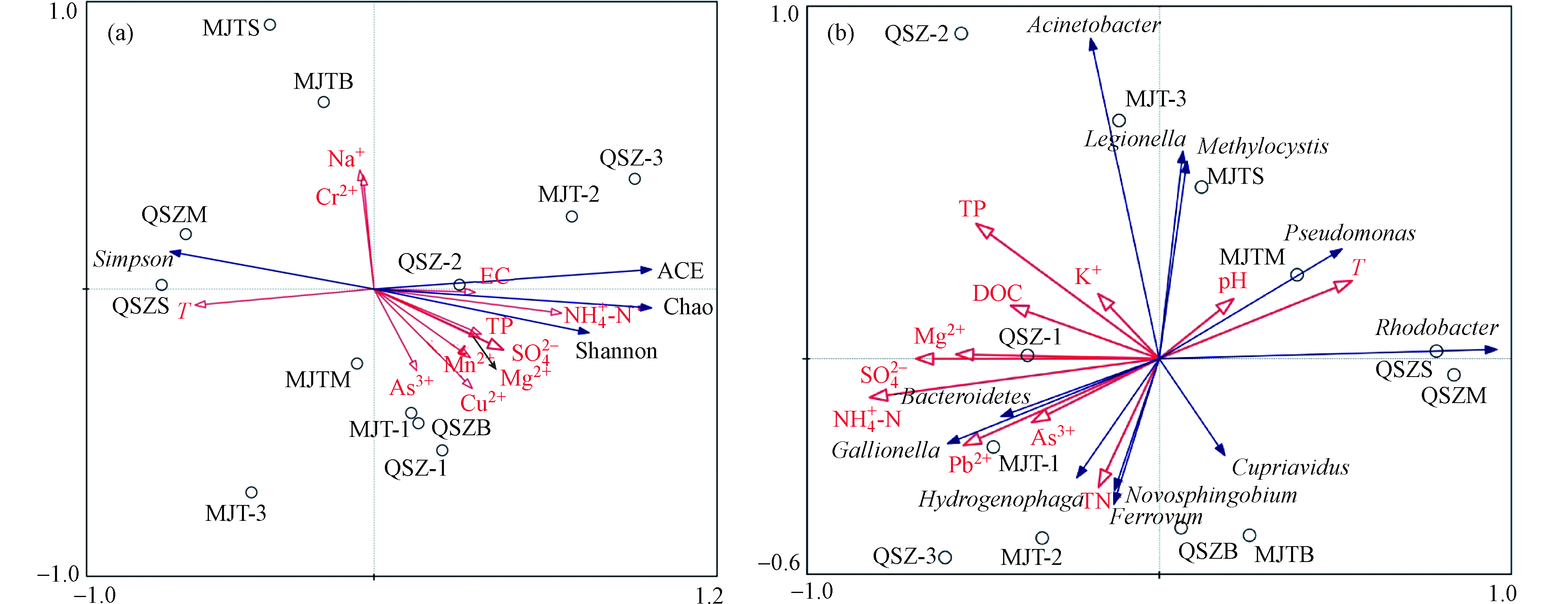
图 4 门水平(a)及属水平(b)细菌群落多样性与环境因子的PCA分析
Figure 4. PCA analysis of bacterial community diversity and environmental factors at phylum level (a) and genus level (b)
表 1 各水库水体主要理化参数
Table 1. Physicochemical parameters of the reservoir water
pH EC/
(μS·cm−1)Eh/
mVDO/
(mg·L−1)T/
℃${\rm{NH}}_4^+ $
(mg·L−1)TN/
(mg·L−1)TP/
(mg·L−1)DOC/
(mg·L−1)${\rm{SO}}_4^{2-} $
(mg·L−1)QSZ-1 3.51 1271.00 460.70 7.44 20.40 0.98 1.80 4.88 2.48 1065.65 QSZ-2 3.74 1106.00 404.50 7.24 21.20 0.86 1.51 5.86 1.13 960.02 QSZ-3 4.02 909.00 382.40 7.19 22.30 0.85 1.54 1.61 1.06 1022.80 QSZS 4.78 598.00 328.80 7.11 23.80 0.07 0.87 0.02 0.69 543.27 QSZM 4.60 580.00 325.10 7.03 23.40 0.13 1.02 0.02 1.00 555.40 QSZB 4.75 557.00 261.50 6.79 23.10 0.59 1.02 0.03 1.38 364.87 MJT-1 3.82 1638.00 417.20 7.14 21.40 0.85 2.11 0.03 1.27 1570.19 MJT-2 6.73 1421.00 -7.10 7.04 21.30 1.06 2.90 0.08 1.39 1112.73 MJT-3 6.56 1132.00 28.60 6.75 24.80 0.93 1.97 0.03 2.24 1042.56 MJTS 6.13 1177.00 174.70 6.84 23.00 0.57 1.02 0.02 0.87 718.66 MJTM 5.70 1211.00 243.30 6.33 22.30 0.35 1.09 0.01 1.04 812.42 MJTB 4.24 1301.00 353.80 5.36 21.80 0.95 5.14 0.04 0.94 832.39 TFe/
(mg·L−1)Mn2+/
(μg·L−1)K+/
(mg·L−1)Na+/
(mg·L−1)Mg2+/
(mg·L−1)Cu2+/
(μg·L−1)Zn2+/
(μg·L−1)Pb2+/
(μg·L−1)As3+/
(μg·L−1)Cr2+/
(μg·L−1)QSZ-1 4.88 3.61 2.98 18.18 62.19 34.11 166.02 43.59 0.63 1.41 QSZ-2 5.86 2.15 3.40 34.24 38.16 21.26 202.32 34.27 0.58 1.58 QSZ-3 1.61 1.70 3.13 27.50 31.02 27.29 286.32 44.85 0.26 2.15 QSZS 0.07 1.12 2.47 15.04 17.28 23.07 415.91 25.93 0.22 1.89 QSZM 0.08 1.16 3.04 15.78 16.65 6.06 77.93 9.19 0.15 0.60 QSZB 1.26 1.13 3.12 15.52 15.78 12.61 110.82 31.65 0.54 0.44 MJT-1 5.10 6.56 4.61 9.50 66.72 43.39 239.20 80.70 5.35 1.84 MJT-2 5.16 3.29 3.34 56.44 61.02 18.69 196.73 83.18 0.59 1.94 MJT-3 1.23 1.46 3.33 59.08 46.71 22.08 119.97 44.81 0.20 1.03 MJTS 0.1 1.83 4.40 69.24 41.16 8.24 88.59 67.66 0.14 0.65 MJTM 0.07 1.89 4.60 69.80 43.80 7.01 74.90 12.67 0.07 0.83 MJTB 0.09 2.06 3.45 75.92 46.44 22.16 107.76 57.23 0.40 16.11  下载: 导出CSV
下载: 导出CSV
表 2 水样浮游细菌多样性统计
Table 2. Diversity statistics of planktonic bacteria in water samples
样品编号
Sample序列数
ReadsOTUs
OUT numbersShannon指数
Shannon indexACE指数
ACE indexChao1指数
Chao1 indexCoverage 指数
Coverage indexSimpson指数
Simpson indexQSZ-1 75559 4646 5.25 7323.34 6842.28 0.97 0.04 QSZ-2 83521 4580 3.36 7783.59 6938.25 0.97 0.18 QSZ-3 110769 6439 4.42 10697.98 9660.64 0.97 0.10 QSZS 97569 1266 1.85 3038.80 2118.58 0.99 0.35 QSZM 86442 1581 2.03 3479.42 2452.01 0.99 0.36 QSZB 64947 4743 5.55 6974.32 6426.88 0.97 0.05 MJT-1 83538 5000 5.21 6871.02 6298.86 0.98 0.03 MJT-2 61293 6444 6.56 9651.25 8682.00 0.96 0.01 MJT-3 85407 2311 2.67 4234.92 3799.55 0.99 0.17 MJTS 95221 2234 2.64 5055.19 3582.34 0.99 0.15 MJTM 90072 3372 3.11 6062.15 5366.32 0.98 0.16 MJTB 82878 3139 4.46 5826.37 4539.25 0.98 0.08
下载: 导出CSV
-
[1] 张河民, 钟铭君, 吴启堂. 石灰石沟-堆肥湿地系统处理酸性矿山废水的研究 [J]. 中国环境科学, 2015, 35(10): 3032-3040. doi: 10.3969/j.issn.1000-6923.2015.10.021 ZHANG H M, ZHONG M J, WU Q T. The treatment of acidic mine drainage using limestone ditch and compost constructed wetland system [J]. China Environmental Science, 2015, 35(10): 3032-3040(in Chinese). doi: 10.3969/j.issn.1000-6923.2015.10.021
[2] GALHARDI J A, BONOTTO D M. Hydrogeochemical features of surface water and groundwater contaminated with acid mine drainage (AMD) in coal mining areas: A case study in southern Brazil [J]. Environmental Science and Pollution Research, 2016, 23(18): 18911-18927. doi: 10.1007/s11356-016-7077-3 [3] KEFENI K K, MSAGATI T A M, MAMBA B B. Acid mine drainage: Prevention, treatment options, and resource recovery: A review [J]. Journal of Cleaner Production, 2017, 151: 475-493. doi: 10.1016/j.jclepro.2017.03.082 [4] 王继勇, 肖挺, 何伟. 一株耐酸 SRB 的分离及其脱硫除镉性能 [J]. 中国环境科学, 2018, 38(11): 4255-4260. doi: 10.3969/j.issn.1000-6923.2018.11.034 WANG J Y, XIAO T, HE W. Isolation and characterization of an acidresistance SRB strain with the efficient of desulfurization and cadmiumremoval [J]. China Environmental Science, 2018, 38(11): 4255-4260(in Chinese). doi: 10.3969/j.issn.1000-6923.2018.11.034
[5] DEAN A P, HARTLEY A, MCINTOSH O A, et al. Metabolic adaptation of a Chlamydomonas acidophila strain isolated from acid mine drainage ponds with low eukaryotic diversity [J]. Science of the Total Environment, 2019, 647: 75-87. doi: 10.1016/j.scitotenv.2018.07.445 [6] 张瑞雪, 吴攀, 杨艳, 等. 贵州煤矿酸性废水“被动处理”技术的新方法探讨 [J]. 地球与环境, 2010, 38(2): 250-254. ZHANG R X, WU P, YANG Y, et al. A new method for passive treatment of acid wastewater from coal mines in Guizhou Province [J]. Earth and environment, 2010, 38(2): 250-254(in Chinese).
[7] 邹莉. 浮游细菌群落结构及氮磷影响因素研究[D]. 北京: 北京大学, 2011. ZOU L. Study on phytoplankton community structure and influencing factors of Nitrogen and phosphorus[D]. Beijing: Peking University, 2011(in Chinese).
[8] 张菲, 田伟, 孙峰, 等. 丹江口库区表层浮游细菌群落组成与PICRUSt功能预测分析 [J]. 环境科学, 2019, 40(3): 1252-1260. ZHANG F, TIAN W, SUN F, et al. Community structure and predictive functional analysis of surface water bacterioplankton in the Danjiangkou Reservoir [J]. Environmental Science, 2019, 40(3): 1252-1260(in Chinese).
[9] 邱琳琳. 黑龙江流域浮游细菌群落组成与环境因子响应关系研究[D]. 哈尔滨: 东北农业大学, 2016. QIU L L. Study on the relationship between phytoplankton community composition and response to environmental factors in Heilongjiang Basin[D]. Harbin: Northeast Agricultural University, 2016(in Chinese).
[10] STALEY C, UNNO T, GOULD T J, et al. Application of Illumina next-generation sequencing to characterize the bacterial community of the Upper Mississippi River [J]. Journal of Applied Microbiology, 2013, 115(5): 1147-1158. doi: 10.1111/jam.12323 [11] FAN L, SONG C, MENG S L, et al. Spatial distribution of planktonic bacterial and archaeal communities in the upper section of the tidal reach in Yangtze River [J]. Scientific Reports, 2016, 6(1): 39147-39147. doi: 10.1038/srep39147 [12] LI Z, LU LH, GUO J S, et al. Responses of spatial-temporal dynamics of bacterioplankton community to large-scale reservoir operation: a case study in the Three Gorges Reservoir, China [J]. Scientific Reports, 2017, 7(1): 42469-42469. doi: 10.1038/srep42469 [13] 沈烽, 赵大勇, 黄睿, 等. 南京玄武湖浮游细菌群落结构的季节变化及其与环境因子的关系[J]. 湖泊科学, 2017, 29(3) : 662-669. SHEN F, ZHAO D Y, HUANG R, et al. Seasonal variation of bacterioplankton community structure in Xuanwu Lake (Nanjing) and its relationship with environmental factors[J]. Lake Sciences, 29(3): 662-669(in Chinese).
[14] 薛银刚, 刘菲, 孙萌, 等. 太湖竺山湾春季浮游细菌群落结构及影响因素 [J]. 环境科学, 2018, 39(3): 1151-1158. XUE Y G, LIU F, SUN M, et al. Community structure and influencing factors of bacterioplankton in spring in Zhushan Bay, Lake Taihu [J]. Environmental Science, 2018, 39(3): 1151-1158(in Chinese).
[15] ZHU C M, ZHANG J Y, NAWAZ M Z, et al. Seasonal succession and spatial distribution of bacterial community structure in a eutrophic freshwater lake, Lake Taihu [J]. Science of the Total Environment, 2019, 669: 29-40. doi: 10.1016/j.scitotenv.2019.03.087 [16] 李俊锋. 基于16S rRNA和宏基因组高通量测序的微生物多样性研究[D]. 北京: 清华大学, 2015. LI J F. Study on microbial diversity based on 16S rRNA and metagenomic high-throughput sequencing[D]. Beijing: Tsinghua University, 2015(in Chinese).
[17] 国家环境保护总局《水和废水监测分析方法》编委会. 水和废水监测分析方法[M]. 第 4 版. 北京: 中国环境科学出版社, 2002 Editorial Committee of "Analytical Methods for Water and Wastewater Monitoring", State Environmental Protection Administration. Analytical methods for water and wastewater monitoring [M]. 4th edition. Beijing: China Environmental Science Press, 2002.
[18] 商潘路, 陈胜男, 黄廷林, 等. 深水型水库热分层诱导水质及真菌种群结构垂向演替 [J]. 环境科学, 2018, 39(3): 1141-1150. SHANG P L, CHEN S N, HUANG Y L, et al. Vertical distribution of fungal community composition and water quality during the deep reservoir thermal stratification [J]. Environmental Science, 2018, 39(3): 1141-1150(in Chinese).
[19] 张广帅, 闫吉顺, 赵全民, 等. 辽东湾小凌河口湿地土壤微生物群落结构与微生态环境因子的关系 [J]. 生态学杂志, 2020, 39(7): 2283-2291. ZHANG G S, YAN J S, ZHAO Q M, et al. Relationship between soil microbial community structure and its micro-ecological environmental factors in Liaodong Bay Xiaolinghe estuarine wetland [J]. Journal of Ecology, 2020, 39(7): 2283-2291(in Chinese).
[20] 邱小琮, 赵红雪, 孙晓雪. 宁夏沙湖浮游植物与水环境因子关系的研究 [J]. 环境科学, 2012, 33(7): 2265-2271. QIU X C, ZHAO H X, SUN X X. Studies on relationship of phytoplankton and water environmental factors in Shahu Lake [J]. Environmental Science, 2012, 33(7): 2265-2271(in Chinese).
[21] HUMAYOUN S B, BANO N, HOLLIBAUGH J T. Depth distribution of microbial diversity in Mono Lake, a Meromictic soda lake in california [J]. Applied and Environmental Microbiology, 2003, 69(2): 1030-1042. doi: 10.1128/AEM.69.2.1030-1042.2003 [22] 范成新. 湖泊沉积物—水界面研究进展与展望 [J]. 湖泊科学, 2019, 31(5): 1191-1218. doi: 10.18307/2019.0514 FANG C X. Research progress and prospect of lake sediment-water interface [J]. Lake science, 2019, 31(5): 1191-1218(in Chinese). doi: 10.18307/2019.0514
[23] LIU Z H, HUANG S B, SUN G P, et al. Phylogenetic diversity, composition and distribution of bacterioplankton community in the Dongjiang River, China [J]. FEMS Microbiology Ecology, 2012, 80(1): 30-44. doi: 10.1111/j.1574-6941.2011.01268.x [24] JACKSON C R, MILLAR J J, PAYNE J T, et al. Free-living and particle-associated bacterioplankton in large rivers of the mississippi river basin demonstrate biogeographic patterns [J]. Applied and Environmental Microbiology, 2014, 80(23): 7186-7195. doi: 10.1128/AEM.01844-14 [25] 王鹏, 陈波, 李传琼, 等. 赣江南昌段丰水期细菌群落特征 [J]. 中国环境科学, 2016, 36(8): 2453-2462. doi: 10.3969/j.issn.1000-6923.2016.08.027 WANG P, CHEN B, LI C Q, et al. Bacterial communities in Nanchang section of the Ganjiang River in wet seaon [J]. China Environmental Science, 2016, 36(8): 2453-2462(in Chinese). doi: 10.3969/j.issn.1000-6923.2016.08.027
[26] 邹沈娟, 尹立强, 赵博礼, 等. 大冶湖浮游细菌群落结构及其环境影响因子研究 [J]. 长江流域资源与环境, 2020, 29(2): 360-366. ZOU S J, YIN L Q, ZHAO B L, et al. Study on phytoplankton community structure and environmental impact factors in Daye Lake [J]. Resources and Environment in the Yangtze Basin, 2020, 29(2): 360-366(in Chinese).
[27] SPRING S, SCHULZE R, OVERMANN J, et al. Identification and characterization of ecologically significant prokaryotes in the sediment of freshwater lakes: molecular and cultivation studies [J]. FEMS Microbiology Reviews, 2000, 24(5): 573-590. doi: 10.1111/j.1574-6976.2000.tb00559.x [28] 杜瑞芳, 李靖宇, 赵吉. 乌梁素海湖滨湿地细菌群落结构多样性 [J]. 微生物学报, 2014, 54(10): 1116-1128. DU R F, LI J Y, ZHAO J. The diversity of bacterial community structure in Suhai Lakeside wetland in Wuliang [J]. Journal of Microbiology, 2014, 54(10): 1116-1128(in Chinese).
[29] TANG X M, GAO G, CHAO J Y, et al. Dynamics of organic-aggregate-associated bacterial communities and related environmental factors in Lake Taihu, a large eutrophic shallow lake in China [J]. Limnology and Oceanography, 2010, 55(2): 469-480. doi: 10.4319/lo.2010.55.2.0469 [30] NEWTON R J, KENT A D, TRIPLETT E W, et al. Microbial community dynamics in a humic lake: differential persistence of common freshwater phylotypes [J]. Environmental Microbiology, 2006, 8(6): 956-970. doi: 10.1111/j.1462-2920.2005.00979.x [31] DISAYATHANOOWAT T, YOSHIYAMA M, KIMURA K, et al. Isolation and characterization of bacteria from the midgut of the Asian honey bee (Apis cerana indica) [J]. Journal of Apicultural Research, 2012, 51(4): 312-319. doi: 10.3896/IBRA.1.51.4.04 [32] BEARDSLEY C, MOSS S, MALFATTI F, et al. Quantitative role of shrimp fecal bacteria in organic matter fluxes in a recirculating shrimp aquaculture system [J]. FEMS Microbiology Ecology, 2011, 77(1): 134-145. doi: 10.1111/j.1574-6941.2011.01094.x [33] ZHANG T, SHAO M F, YE L. 454 pyrosequencing reveals bacterial diversityof activated sludge from 14 sewage treatment plants [J]. The ISME Journal, 2012, 6(6): 1137-1147. doi: 10.1038/ismej.2011.188 [34] 司开学, 夏长革, 王朝阳, 等. 宁波沿海陆源排污口放线菌(Actinomycetales sp. ) 的分布特点 [J]. 海洋与湖沼, 2016, 47(2): 400-406. SI K X, XIA C G, WANG C Y, et al. Distribution of Actinomycetales sp. in sewage outlets along coast in Ningbo [J]. Oceans and Lakes, 2016, 47(2): 400-406(in Chinese).
[35] JETTEN M S M. The microbial nitrogen cycle [J]. Environmental Microbiology, 2008, 10(1): 2903-2909. [36] 白洁, 李海艳, 赵阳国. 黄海北部不同站位海洋细菌群落分布特征 [J]. 微生物学报, 2009, 49(3): 343-350. doi: 10.3321/j.issn:0001-6209.2009.03.010 BAI J, LI H Y, ZHAO Y G. Distribution characteristics of Marine bacterial community at different stations in the northern Yellow Sea [J]. Journal of Microbiology, 2009, 49(3): 343-350(in Chinese). doi: 10.3321/j.issn:0001-6209.2009.03.010
[37] DWORKIN M, FALKOW S, ROSENBERG E, et al. The prokaryotes[M]. New York: Springer, 2006. [38] WANG L, ZHANG J, LI H L, et al. Shift in the microbial community composition of surface water and sediment along an urban river [J]. Science of the Total Environment, 2018, 627: 600-612. doi: 10.1016/j.scitotenv.2018.01.203 [39] MCLELLAN S L, NEWTON R J, VANDEWALLE J L, et al. Sewage reflects the distribution of human faecal lachnospiraceae [J]. Environmental Microbiology, 2013, 15(8): 2213-2227. doi: 10.1111/1462-2920.12092 [40] 罗宁, 罗固源, 吉方英, 等. 利用聚磷菌快速内源性反硝化脱氮研究 [J]. 三峡环境与生态, 2003, 25(6): 32-34,58. LUO N, LUO G Y, JI F Y, et al. Rapid endogenous denitrification with polyphosphobacteria was studied [J]. Environment and Ecology of the Three Gorges, 2003, 25(6): 32-34,58(in Chinese).
[41] ROGUET A, LAIGLE G S, THERIAL C, et al. Neutral community model explains the bacterial community assembly in freshwater lakes[J]. FEMS Microbiology Ecology, 2015, 91(11),DOI: 10.1093/femsec/fiv125. [42] WANG S R, ZHAO D Y, ZENG J, et al. Variations of bacterial community during the decomposition of microcystis under different temperatures and biomass [J]. BMC Microbiology, 2019, 19(1): 1-10. doi: 10.1186/s12866-018-1372-8 [43] 蔡林林, 周巧红, 王川. 南淝河细菌群落结构的研究 [J]. 环境科学与技术, 2012, 35(3): 1-6. doi: 10.3969/j.issn.1003-6504.2012.03.001 CAI L L, ZHOU Q H, WANG C. Study on the bacterial community structure of Nanfeihe River [J]. Environmental Science and Technology, 2012, 35(3): 1-6(in Chinese). doi: 10.3969/j.issn.1003-6504.2012.03.001
[44] 李玉华, 许其功, 赵越, 等. 松花湖水体中不同空间分布的细菌群落结构分析 [J]. 农业环境科学学报, 2013, 32(4): 764-770. LI Y H, XU Q G, ZHAO Y, et al. Bacterial community structure in different spatial distribution of Songhua Lake [J]. Journal of Agro-Environment Science, 2013, 32(4): 764-770(in Chinese).
[45] GUILDFORD S J, HECKY R E. Total nitrogen, total phosphorus, and nutrient limitation in lakes and oceans: Is there a common relationship? [J]. Limnology and Oceanography, 2000, 45(6): 1213-1223. doi: 10.4319/lo.2000.45.6.1213 [46] SCHULTZE M, POKRANDT K, HILLE W. Pit lakes of the central german lignite mining district: Creation, morphometry and water quality aspects [J]. Limnologica, 2010, 40(2): 148-155. doi: 10.1016/j.limno.2009.11.006 [47] JESCHKE C, FALAGAN C, KNOLLER K, et al. No nitrification in lakes below pH3 [J]. Environmental Science & Technology, 2013, 47(24): 14018-14023. [48] ZHANG L, SHEN T T, CHENG Y, et al. Temporal and spatial variations in the bacterial community composition in Lake Bosten, a large, brackish lake in China [J]. Scientific reports, 2020, 10(1): 1-10. doi: 10.1038/s41598-019-56847-4 [49] DAS B K, ROY A, KOSCHORRECK M, et al. Occurrence and role of algae and fungi in acid mine drainage environment with special reference to metals and sulfate immobilization [J]. Water Research, 2009, 43(4): 883-894. doi: 10.1016/j.watres.2008.11.046 -
 点击查看大图
点击查看大图
计量
- 文章访问数: 3264
- HTML全文浏览数: 3264
- PDF下载数: 92
- 施引文献: 0


















