



-
酸雨作为工业过程伴生的环境问题,关乎全球环境. 作为酸雨污染大国,我国近40%面积受酸雨污染,对农业生产危害不容忽视[1-2]. 酸雨作用于叶面,诱发H+与叶片中阳离子竞争结合位点与离子替换,致细胞酸化坏死,酸雨中SO2经气孔对植株产生伤害[3-5]. 细胞内酸性物质累积使线粒体和叶绿体超微结构改变,导致叶绿素功能异常,呼吸及光合作用减弱[6];酸雨胁迫还引发活性氧累积,引起细胞膜脂质过氧化损伤[7],抑制抗氧化酶基因表达及活性[8]. 种子萌发和苗期生理生化指标受酸雨影响巨大[7]. 酸雨导致土壤酸化,使土壤中铝、镉等有害元素析出,抑制根系活性[9-10]、养分吸收、代谢及生长. 土壤酸化进一步造成微生物多样性[10]及活性降低[8],阻滞植物生长发育. 大量研究表明,玉米[11-12]、水稻[13]、小麦[14]其生长发育过程中物质转运吸收效率在pH低于5的条件下会明显受阻,当pH进一步降低到3以下时,多种作物的生理生态过程将出现停滞的现象[12]. 随着全球人口激增,酸雨威胁粮食生产安全. 人们通过农药及化肥增强作物抗性以期产量增加的努力,对环境和人体健康存在隐患. 因此,在当下无法控制酸雨污染、培育抗逆(酸雨)植株的现实条件下,寻找一种低污染、见效快、易推广的化控减灾技术与产品,迫在眉睫.
农业上稀土元素(rare earth elements, REE)镧使用广泛[15-17]. 稀土施用水稻[13]、油菜[12]和黄瓜幼苗[18],其处理组氮、磷、钾等含量远高于对照组,生长更优. 油菜叶绿素含量、硝酸还原酶活性及根系活力、产量增效明显[12]. REE提高糯玉米幼苗PSⅡ最大量子产额和电子传递率,增强光合作用与抗氧化酶活性(减少丙二醛累积)及幼苗抗逆性[19]. 众多实践证明,La3+能提高玉米、苏丹草、紫花苜蓿[20-22]抗盐碱能力,降低玉米和小麦[14, 20]叶片蒸腾速率,增强植株抗逆性. 作为植物成分的氨基酸,既是植物生存的养分,也可凭其络合能力对植物的生理生态形成调节作用[23]. 如氨基酸浸种能缩短水稻与高粱种子萌发时间,提高淀粉酶活性与种子活力[24]. 甘氨酸缩短小油菜生育周期[24],提高烟草叶面积[25]、叶绿素含量及光合作用[25]. 色氨酸增加草莓叶厚(密)度,促进生长发育[26]. 脯氨酸能提高水稻细胞抗氧化酶活性及叶绿素和类胡萝卜素含量,减少丙二醛累积,增加植株抗逆性及生物量[27];氨基丁酸能提高烟草抗氧化酶活性,缓解盐碱对生长发育胁迫[25]. 谷氨酸提高严寒中小白菜叶含水量、鲜重、光合效率及对低温耐受性,甘氨酸增加棉花过氧化物酶等活,控制细胞活性氧水平,增强植株抗逆性[28].
稀土-氨基酸络合物具优良性能、稳定结构、原料获取便捷、合成过程精炼,较单独施用更有优势,其以较高的性价比已成为提高植物抗逆性、用途广泛的产品之一. Zhong等[29]发现,镧螯合氨基酸能缓解铜离子对水稻胁迫;周芸[30]在研究氨基酸稀土微肥时也看到,它能有效提高花生果实干物质累积、果实密度及出仁率,促进增产. 遗憾的是,有关特定稀土螯合氨基酸用于作物化控减灾的研究尚少.
本文采用模拟酸雨(pH3.0)处理玉米(Zea mays L.)、水稻(Oryza sativa L.)、小麦(Triticum aestivum L.)等3种作物的实验设计,运用生理生化等技术手段,初步研究新型氨基酸螯合镧(La(Ⅲ)-AA)对酸雨胁迫下植物质膜透性(MP)、抗氧化系统(过氧化物酶POD、过氧化氢酶CAT、超氧阴离子自由基ROS产生速率、丙二醛MDA含量)、光合作用能力(叶绿素含量)、脯氨酸(PRO)含量及氮代谢(硝酸还原酶NR)等生理指标影响,为酸雨胁迫下作物化控减灾,以及调控产品的选择提供依据.
-
参照ZHONG等人[29]的研究成果,在60 ℃及pH8的条件下以10%碱性蛋白酶水解大豆分离蛋白(SPI) 2 h,随后经100 ℃高温灭活10 min,取上清液在4 ℃、10000 r·min−1下离心10 min. 50 ℃条件将上述制备液与La (NO3)3·6H2O以1.5∶1的体积比反应5 h制得螯合液. 将螯合液置于4 ℃、10000 r·min−1下离心10 min,上清液用丙酮沉淀干燥,最终得到氨基酸螯合镧(La(Ⅲ)-AA)供试品.
经过前人研究可知,植物体生长发育过程中其表面叶绿素含量与逆境情况呈现负相关[31],且在酸雨胁迫过程中植物体内叶绿素含量会出现明显的降低,因此本次研究综合参照Zhong等[29]有关La(Ⅲ)-AA对Cu2+胁迫研究以及前期预实验针对La(Ⅲ)-AA对3种作物的喷施后叶绿素表现结果综合研判,确定20 mg·L−1的施用浓度作为本轮研究的施用浓度.
-
模拟酸雨配制参考江南地区酸雨降水条件,并参照陈俊彤等[4]有关茼蒿的研究和实验用书方案[32]进行配制,其比例为
${\rm{SO}}_4^{2-} $ :${\rm{NO}}_3^{-} $ =4.7:1(V:V). 实验过程中先配制pH为1.0的酸雨储备液备用,后续施加过程中再依据研究需求与少量磷酸盐缓冲液混合制成pH为3.0的模拟酸雨,储备液及模拟酸雨的制备过程均采用pHS-29A酸度计进行pH校准. -
玉米(Zea mays L.)、水稻(Oryza sativa L.)、小麦(Triticum aestivum L.)由相城区种苗繁育基地提供,选取苗种茁壮,高度约为60 cm,5—6叶玉米(Zea mays L.)幼苗、高度约为45 cm,4—5叶水稻(Oryza sativa L.)幼苗和高度约为35 cm,2—3节小麦(Triticum aestivum L.)幼苗的种苗移栽,随后在实验室预栽培5—10 d以适应新的生长实验条件,选取繁育室温度控制在(25±2) ℃[32],以T5型全光谱组培灯作为辐射光源(OPPLE、T5-1058-18 W),光子通量密度设置为2000 μmol·m−2·s−1,光照距离为15 cm,并随幼苗生长而随时调整辐射光源高度[4, 19, 32]. 实验过程中,光暗比为9 h/15 h[32],每3 d进行一次换水,每日早晚各通气1次,实验过程均采用Hoagland培养液水基培养以满足植株幼苗的生长需求[30]. 实验过程中不同组别的设置方式如表1中所示.
为增加喷施溶液在植物叶片表面的沾附能力,在蒸馏水中添加3—5滴聚山梨酯-80,La(Ⅲ)-AA 20 mg·L−1溶液及酸雨处理液中添加1—2滴为宜[4,33],实验周期内植株幼苗均栽种于15穴育苗盘(高度×边长为98 mm×75 mm)中,每穴1株幼苗以便处理.
供试植株经表1方式处理后培养24 h开始计算实验时间,随后在实验时间第1、5、10、15 天对3种供试植株幼苗的组织进行生理指标测定,为减少人为测定误差,每个时间节点每个品种均随机选取3株进行测定,单一样本进行5次测定取平均值作为实验数据.
-
质膜透性测定(MP),采用电导率法[34];过氧化物酶(POD)、超氧阴离子自由基产生速率(ROS)以及硝酸还原酶(NR)活性,其测定方法参照植物生理学实验指导(第五版)执行[32];丙二醛(MDA)含量,采用硫代巴比妥酸法测定[3];叶绿素含量,通过SPAD502手持式叶绿素仪直接测定法来表示[35];脯氨酸含量(PRO),采用磺基水杨酸-茚三酮法[35].
-
研究中数据分析统计均采用SPSS 22.0软件进行,平均值、标准差和相对值,单因素方差分析差异性,采用最小显著性差数法(LSD),即以P<0.05作为差异显著的判断标准,图表采用Origin Pro 2017进行绘制.
-
细胞质膜是分隔细胞内外环境屏障,用以维持细胞微环境稳态. 图1显示,随酸雨胁迫时间延长,玉米、水稻、小麦幼苗质膜透性较对照组(BK)增加(小麦15 d除外);预喷施La(Ⅲ)-AA组(BS)的3种作物,质膜透性增幅弱于酸雨组(玉米1 d除外),甚至低于BK组(玉米10 d、15 d,水稻10 d、15 d,小麦组5 d、10 d、15 d);BS组相对于AR组则是呈现降低态势(玉米1 d除外),其差异极值分别出现在10 d(玉米)、10 d(水稻)、10 d(小麦),BS组相对电导率仅为AR组的51.42%、54.22%和45.95%. 其质膜透性结果与姚梦婕等[34]对稀土铈对酸雨胁迫过程中水稻幼苗的防护具有相似的情况,表明质膜是非生物胁迫过程中逆境作用靶位,大量自由基攻击膜脂不饱和脂肪酸双键,细胞质膜氧化分解受损[4],失去离子筛选功能,进而导致大量有害离子和游离基团涌入胞内形成强大破坏力,最终造成加剧细胞膜脂过氧化,对质膜形成更严重破坏,植株生理功能不断恶化[5]. 酸雨胁迫下,质膜透性与相对电导率增加,植物受害加重,抗逆性减弱[36]. 而随着La(Ⅲ)-AA的施加其在植物叶片表面形成保护膜,氨基酸成分在提供营养物质的同时对酸性环境起到了良好的缓冲作用,进而对质膜起到良好保护 [4, 7],从而减少外界H+、SO2等有害物质输入诱发的植株体自由基成分的代谢失衡. La(Ⅲ)-AA所表现出的缓冲作用与冯宗炜[2]早期所提出的提高土壤缓冲能力进而防治酸雨的对策具有异曲同工之妙. 上述结果可知,酸雨胁迫使作物膜损伤、质膜透性增加. La(Ⅲ)-AA可明显增强植株细胞质膜稳定性,降低质膜透性,从而减轻酸雨胁迫对植株造成伤害.
-
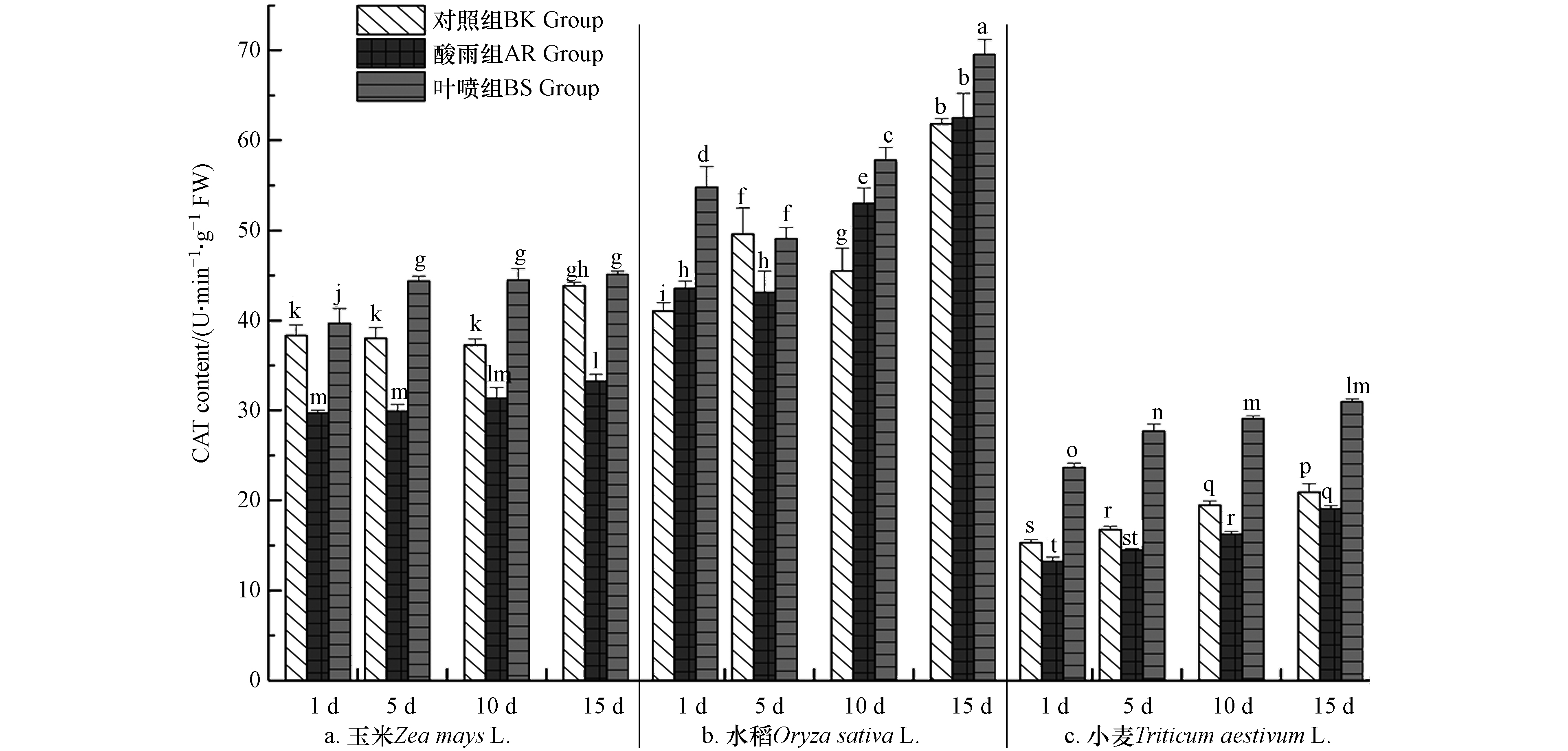
CAT酶能清除植物体内过氧化氢,防止质膜过氧化损伤,延缓植物衰老. 图2中数据直观的表明,小麦CAT活性相较于玉米和水稻,总体偏低,其酶活最高点也仅为玉米种BK组最低酶活的80.51%;植株体内CAT活性AR组低于BK组(水稻1 d、10 d、15 d除外),表明酸雨对于作物CAT酶活性有着抑制,且随着时间延长,系统的CAT酶活均出现了增长(水稻10 d除外);BS组其CAT活性相较于BK组有较为显著的提升,说明La(Ⅲ)-AA对作物体内CAT酶活有促进作用;而BS组酶活均高于AR组,且在3种作物中的差异极值分别出现在5 d(玉米)、1 d(水稻)、10 d(小麦),La(Ⅲ)-AA使得BS组相较于AR组种CAT活性显著提高了48.68%、29.43%、78.61%,结果表明酸雨使得作物体CAT酶活性受抑. 而La(Ⅲ)-AA 能够很好的消除酸雨对CAT酶的抑制作用,且使得CAT酶活大幅度提高,有效地对植株体内产生的过氧化氢起到应答作用,将植物体内过氧化氢含量维持在较为稳定的水平,更好地为作物生长发育提供保障.
-
POD酶活性及同工酶表达形式与植物生长发育过程相关,其酶活随植株老化不断升高[4]. 图3数据可知,POD酶活在不同作物中呈现出较大差异,其在玉米植株中活性表现最低而在小麦中活性最高;单一植株组内数据比对不难发现,POD活性随着时间的延长总体呈现出下降的趋势,且酸雨胁迫下植株体内POD酶活性显著升高(除小麦1 d);La(Ⅲ)-AA则有效降低了酸雨胁迫下POD酶活,且在1 d(玉米)、10 d(水稻)、15 d(小麦)时使得其与AR组相差最大,分别为AR组的48.3%、57.92%、50.44%,以至于玉米1 d、10 d、15 d,水稻5 d、10 d,小麦5 d、10 d、15 d时,其POD活性低于BK组. 上述结果表明,酸雨胁迫下植株内过氧化物及游离自由基含量逐步增多,加速植株体的老化. 而La(Ⅲ)-AA有效地为植株提供了防护作用,减少其机体内过氧化物产生量,进而表现为POD含量大幅度降低,由此可见其某种程度上不仅能够有效地缓解酸雨胁迫对作物机体的破坏,而且对植物生理发育具有促进作用.
植物体中两大标志性抗氧化酶系统(CAT和POD)的结果变化表明,植物在系统演化过程中,构建出修复逆境伤害的机制[13]. 酸雨胁迫下,作物自身应答机制发挥作用,体内抗氧化物酶协同作用,维持细胞稳态. 此轮研究结果中AR组CAT活性在出现显著低于BK组的现象,其与唐加红等[14]对小麦抗旱的研究有所出入,小麦在干旱胁迫下其体内的三大抗氧化酶系均呈现出明显的增长. 推测造成此次变化的主要原因是由于POD作用于不同底物,在氧化底物同时将氧还原成H2O2,后者被CAT再次分解(CAT活性增长存在一定延迟性),因此本轮研究中CAT酶活随时间延续呈渐增趋势(图2),而POD酶活增幅则明显趋前(图3). 而此种CAT略微延迟的现象在任鹏辉[22]对紫花苜蓿的抗盐胁迫过程中得以呈现. 植株通过CAT及POD酶活变化,不断调节自身机能,从而更好适应酸雨胁迫,为植株生长发育提供保障. 而通过上述BS组与AR组的数据分析结果可以了解到,La(Ⅲ)-AA能够有效缓解酸雨胁迫对抗氧化酶所产生的影响,因此La(Ⅲ)-AA对植物体抗氧化酶系起到了积极的促进作用.
-
逆境条件下,植物产生较多ROS给机体造成不可逆转伤害,抑制植物长发育[22]. 玉米、水稻、小麦作物体内ROS含量如图4所示,由于物种差异ROS产生速率在玉米中远高于水稻和小麦;酸雨胁迫下AR组ROS产生速率相较BK组均有显著的增长;BS组在La(Ⅲ)-AA 作用下ROS速率得以降低,且均低于AR组和BK组(小麦10 d除外),BS组相较于AR组,在10 d(玉米)、1 d(水稻)、15 d(小麦)ROS降低最为明显,分别降低了28.44%、39.47%、42.31%. 以上数据得以证明酸雨对植株体的生长发育产生了较为严重的胁迫作用,其植株体的质膜受损从而加大其体内ROS的不断升高和累积,如不加以控制最终会造成不可逆的伤害,其与金琎等[11]对玉米的酸雨抗性研究具有相同的结果趋势. 而随着La(Ⅲ)-AA的施用其ROS速率得到了较为有效的抑制,其不仅能够缓解酸雨胁迫给植物体所带来的危害,而且能够对植株体生长发育环境形成优良的缓冲,减少植物所受到的环境胁迫影响.
-
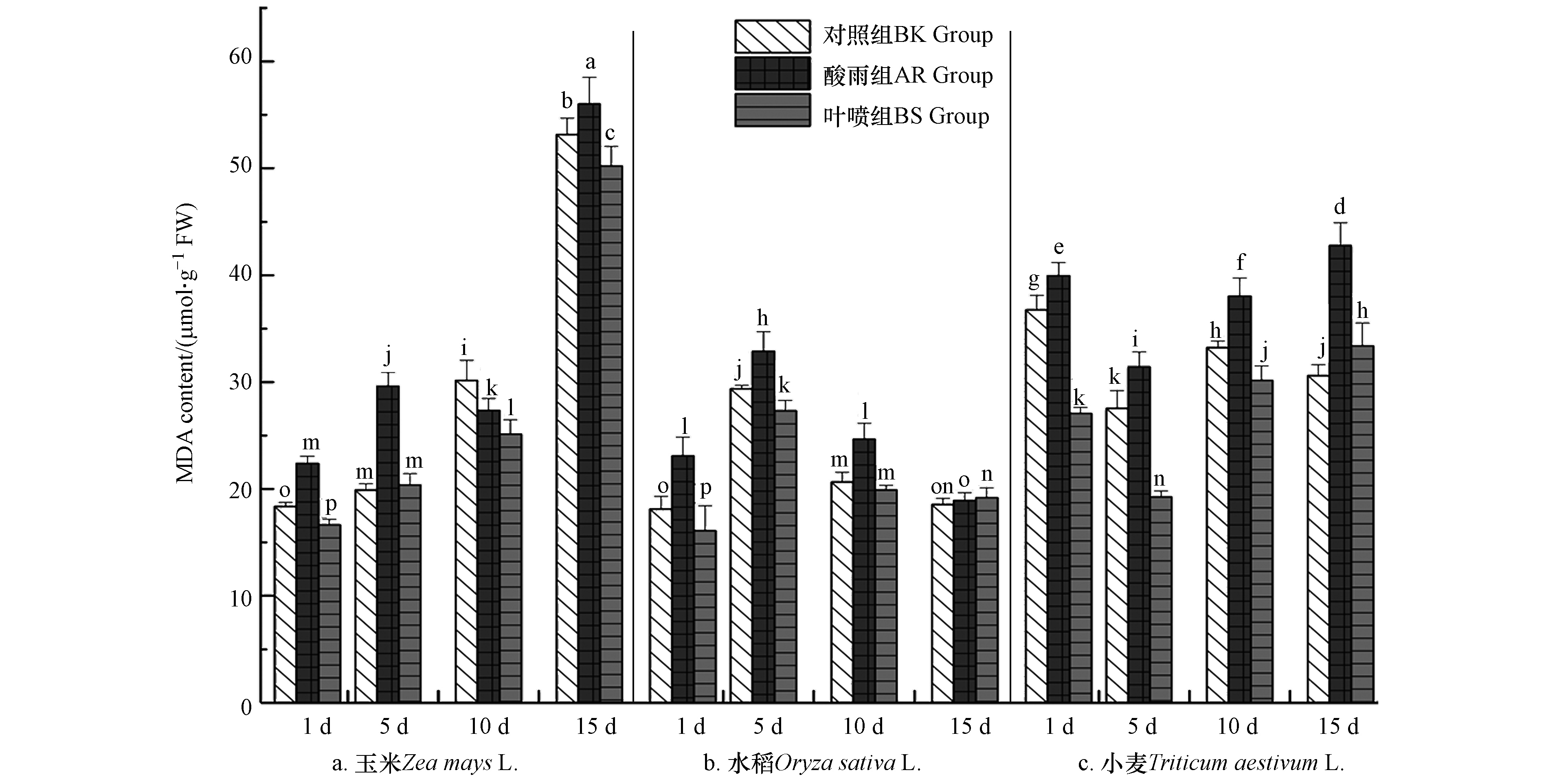
植株体过氧化最直接的产物便是MDA,其能够有效地反映植株体当前的健康水平[4]. 图5中数据可以清晰的观察到,玉米和小麦作物中MDA的水平随着培育时间的延长总体呈现上升的趋势,而水稻种的MDA则在5 d达到最高,随后逐步回落;随着酸雨对作物胁迫时间的逐步延长,玉米和小麦中的MDA均出现持续的累积,这两种作物MDA的最高值均出现在15 d的AR组,而水稻作物中的MDA在5 d后呈现稳步回落,直至15 d其组内3种处理方式作物中MDA含量接近,AR组及BS组回归到正常水平;除水稻15 d外,BS组中MDA的含量相较于AR组均有着显著的降低,其降低的峰值在5 d(玉米)、1 d(水稻)和5 d(小麦),分别降低了31.03%、34.78%、38.71%. 其结果与巩东辉等[16]对螺旋藻的生理生态指标研究具有一定的相似性. 在外界环境胁迫下细胞质膜失去了其原本的防护作用, ROS的大量增加加剧其胞内物质的过氧化,其过氧化产物MDA便呈现出增长趋势,进而其反作用于细胞质膜,亦是对其形成更严重破坏,植株生理功能不断恶化[5]. 由此可见,酸雨会加速MDA在植株体内的累积进而对植株体产生破坏作用. 而La(Ⅲ)-AA对酸雨胁迫下植物中MDA的产生具有缓解作用,能够较为有效的减少植物机体损伤.
-
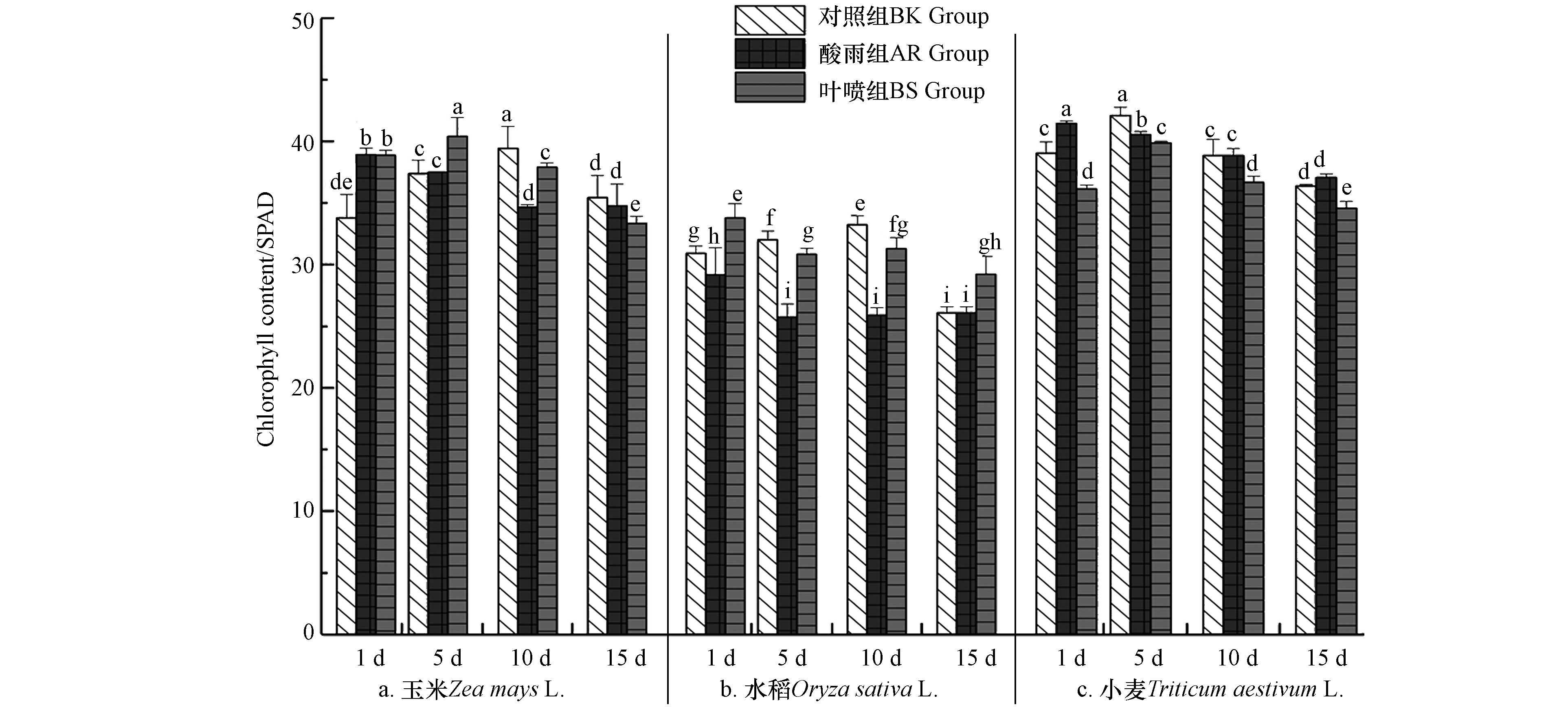
叶绿素是植物体光合作用及碳代谢的关键节点,其存在对植物体生长发育起到决定性作用. 图6可以了解到,叶绿素在3种作物叶片中的含量随着培育时间的延长总体呈现先扬后抑的趋势,BK组叶绿素最高点分别出现在10 d(玉米)、10 d(水稻)、5 d(小麦);酸雨的环境胁迫情况总体在水稻种体现的更为明显,其AR组的叶绿素含量均显著低于BK组,而在玉米和小麦体内则出现了分化,玉米组1 d时AR组叶绿素含量高于BK组14.71%,与此同时小麦组1 d和15 d中亦是出现AR组高于BK组的情况;而BS组中水稻和玉米作物的叶绿素含量相较AR组均有一定幅度的增长,水稻中为18.76%(10 d),玉米中为4.21%(10 d),而小麦作物中La(Ⅲ)-AA对其叶绿素含量反而会有降低的影响. 造成小麦作物中La(Ⅲ)-AA表现不佳的因素与李月福等[12]研究中油菜幼苗的应激过程具有一定的相似性,La(Ⅲ)的对植株叶绿素的改观效果与植株本体所受到的胁迫程度具有一定的相关性,其在不同胁迫状况下La(Ⅲ)的左右效果往往具有一定差异,此时适当调整降低La(Ⅲ)浓度便可获得较优的效果. 而酸雨胁迫过程中叶片细胞叶绿体结构受害,色素合成途径受阻,叶绿素合成量减少并伴随叶绿素降解[22],随胁迫时间延长,叶绿素含量渐降(图6)并因此影响植株体生理活性. 而La(Ⅲ)-AA能够在叶表面进入叶细胞后,供植物生长所需微量元素和必需氨基酸,从而缓解酸雨污染对细胞微环境扰动. 或将叶绿素中心的Mg取代,进而形成夹心螯合物,在提高叶绿素稳定性同时,也使其捕获波长较短能量较高光子的能力大幅提高,有助于光合效率提升[33],增强其对CO2催化能力,提高机体内糖类物质累积. 综合数据表明,不同植株体对酸雨和La(Ⅲ)-AA的作用不尽相同,但总体来说La(Ⅲ)-AA能够有效缓解酸雨所带来的影响.
-
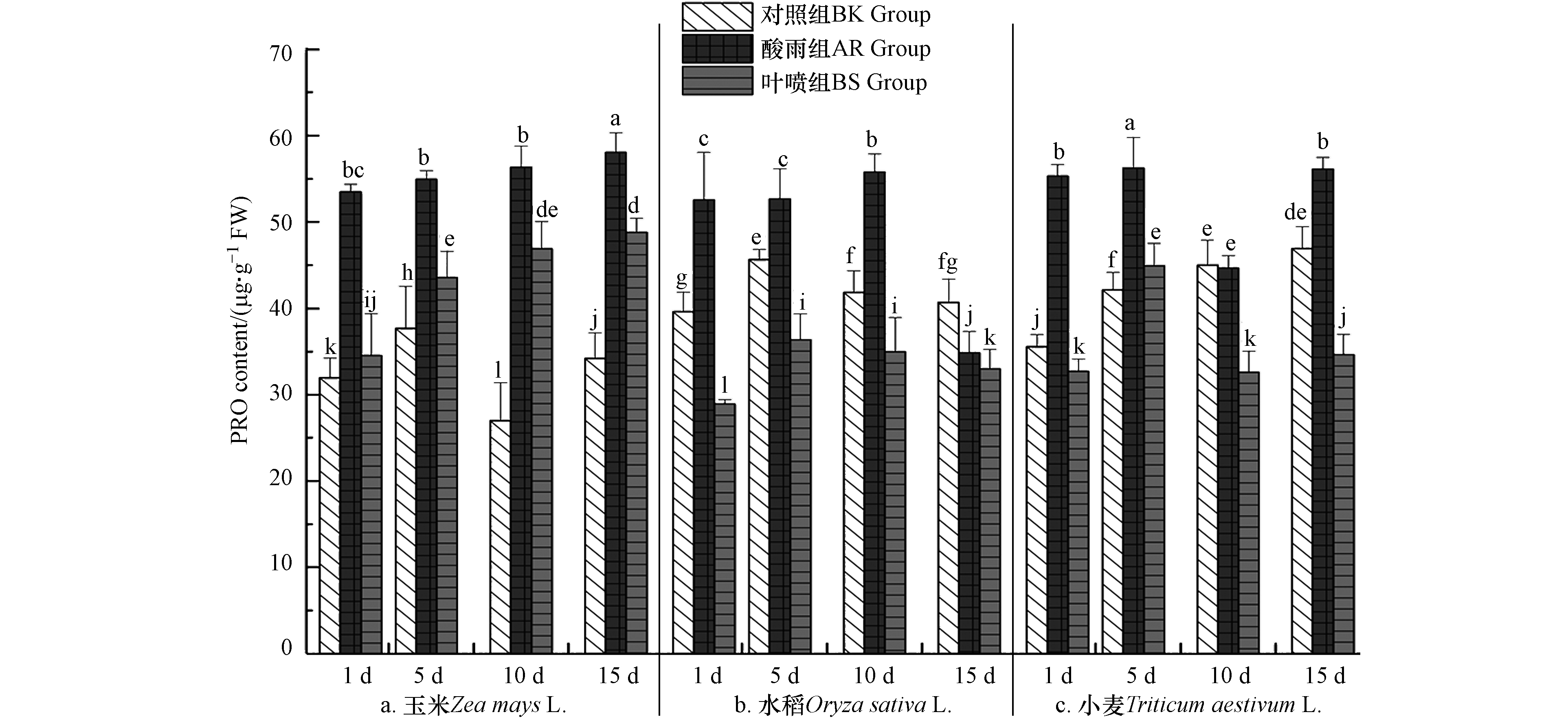
游离态PRO在构成植物蛋白的同时在维持渗透压、稳定分子结构、调节细胞氧化过程方面亦是具有重要作用[4]. PRO的改变情况如图7中所示,PRO在不同植物体中的含量总体趋于平稳,3组作物中BK组中PRO均在40 μg·g−1FW上下浮动;酸雨胁迫使得植物体做出对应的应答反应,3种作物中AR组的PRO含量均出现了显著的提高;而BS组中PRO含量相较于AR组均大幅度降低,3种作物中均在1 d出现最大的减少幅度,分别为36.23%(玉米)、47.17%(水稻)、41.82%(小麦),且在玉米和水稻中多次出现BS组低于BK组的现象. PRO并不是独立存在的个体,其在植株体受到外界环境胁迫的过程中会进行大量的累积[5],体内PRO与抗氧化物酶协同作用,维持细胞稳态. 即当亲水性强的PRO含量升高、游离态PRO广泛存在时,为稳定细胞内大分子蛋白,降低功能蛋白酸解及渗透压调节和平衡等代谢过程[7]提供保障[12]. 上述结果表明,酸雨对植株体生理生化产生了严重的干扰和破坏,PRO在植物体内大量的积累以稳定植物体细胞所带来的渗透压从而为植株更好的生长提供必要条件. 而La(Ⅲ)-AA为作物在酸雨胁迫下提供了优良的缓冲作用,使得酸雨危害大大的减少,为植物体抗逆性提供坚实的保障.
-
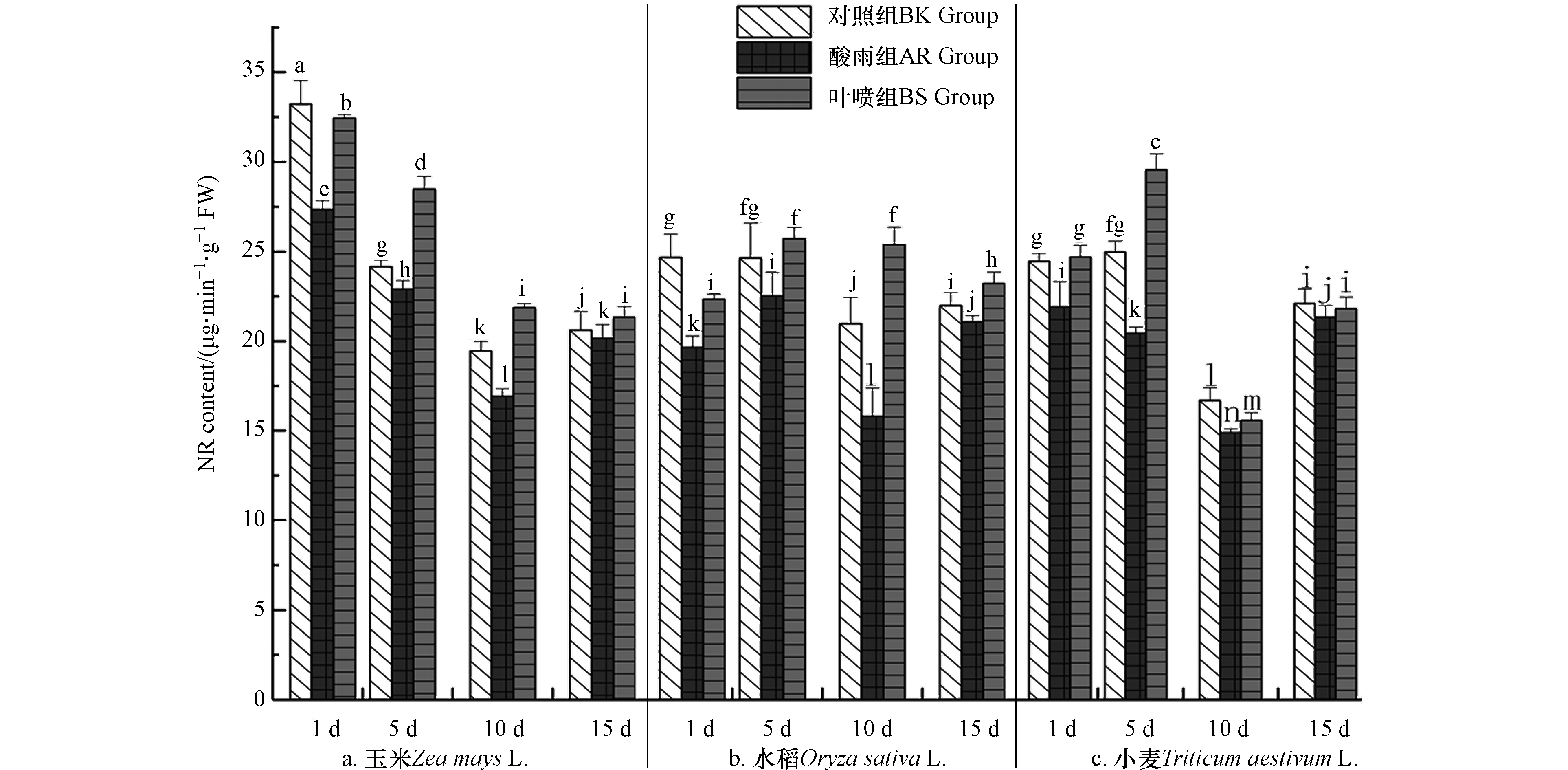
硝酸还原酶(NR)是N元素有效利用的关键酶,其能够较直观地反映出植物体N代谢的强度[32]. NR活性如图7中所示,总体看来3种作物中NR活性呈现出先抑后扬的趋势,BK组均在10 d出现最低值,随后升高;AR组中NR活性均出现了不同程度降低;BS组NR活性具有大幅度提高,玉米(5 d、10 d、15 d)、水稻(5 d、10 d、15 d)、小麦(1 d、5 d)多次高于BK组,相较于AR组,在5 d(玉米)、10 d(水稻)、5 d(小麦)NR增幅最为显著,分别增长了21.37%、60.63%、46.50%,与AR组形成鲜明的对比. 酸雨胁迫下对叶绿素的影响逐渐传递并表现为到氮代谢过程的干扰,叶绿素含量渐降(图6)并因此影响NR酶活. 这是因为,NR酶是以细胞色素作为电子供体[35],色素合成受阻引起NR酶催化反应减弱(图8),细胞内NO3-还原成NO2-速率降低,降低植物氮代谢水平. 且亚硝酸盐转化NO是多种代谢途径的信号物质,如根系生长、营养物质吸收及有机酸代谢[18]. 植物氮代谢水平降低,影响其生长发育,降低抗逆性,最终表现为酸雨对3种作物伤害.
而La(Ⅲ)-AA 施用后可以使得ROS进一步减少,与此同时保持了细胞微环境稳态,其最直接的结果便是,细胞叶绿体微结构、叶绿素含量与细胞色素电子传递链功能趋于正常,以满足NR酶促反应对光电子需求[30],确保NR酶活与细胞内NO3-还原成NO2-速率,维持植物氮代谢水平,以及信号物质NO参与的多种代谢途径调控[33]. 植物氮代谢正常对其生长发育、抗逆性不无裨益,并最终促进植物更好生长发育,产量提高.
上述数据不难看出,酸雨对于植物体氮代谢具有较大的抑制作用,不利于植物生长. 而La(Ⅲ)-AA能够显著提升NR活性,增强植株体对酸雨胁迫的抗性,且其在一定程度上能够促进作物本身的氮代谢强度,提高物质积累,进而对作物生长发育过程形成增益.
实验中还观察到,3种作物各项生理生化指标对酸雨胁迫强度、时长存在差异反应(图1—8),根据已有报道推测,或与3种作物对酸雨胁迫的敏感性有关. 如在耐酸性的生态适应上,水稻耐酸性强于小麦、玉米[37-38]. 当然,更准确的答案也许要在酸雨胁迫下,植物逆境基因确认与响应上寻求.
-
(1)酸雨胁迫对3中经济作物造成显著损伤,作物体内MP、ROS产生速率、MDA含量、PRO含量、POD活性均显著升高. 与此同时,叶绿素含量、CAT活性、NR活性则显著降低;
(2)预施La(Ⅲ)-AA溶液,可有效缓解酸雨对作物产生的胁迫伤害,MP、ROS产生速率、MDA含量、PRO含量降低,CAT和NR活性提高,POD活性趋于稳定;
(3)预施La(Ⅲ)-AA产生防护效果,为化控酸雨损害作物的研究提供新的可行性.
氨基酸螯合镧(La(Ⅲ)-AA)对模拟酸雨胁迫下三种作物生理指标的影响
Effects of La(Ⅲ)-AA on Physiological Indicators of three crops under artificial acid rain stress
-
摘要: 酸雨对农业生态系统中作物的影响,攸关粮食安全,抗酸(雨)减灾研究迫在眉睫. 本文采用模拟酸雨污染盆栽实验方法,研究一种新型氨基酸螯合镧对酸雨胁迫下玉米、水稻、小麦等8项生理指标的影响. 结果表明,与对照组相比预施氨基酸螯合镧能有效降低酸雨对3种作物叶片质膜透性(45.95%—54.22%)、超氧阴离子自由基产生速率(28.44%—42.31%)、丙二醛含量(31.03%—38.71%)、脯氨酸(36.23%—47.17%)含量及过氧化物酶活性(48.3%—57.92%)的影响,提高过氧化氢酶活性(29.43%—78.61%)、叶绿素含量(7.89%—18.76%)和氮代谢强度(21.37%—60.63%). 由此可见,氨基酸螯合镧对酸雨胁迫下,作物的过氧化损伤具有一定防护效应,预喷氨基酸螯合镧可在一定程度上减轻酸雨污染对作物的伤害.Abstract: The impact of acid rain on crops is critical to food security, and reducing acid rain damage to crops is a top priority. In this paper, the effect of a novel amino acid chelated lanthanum La(Ⅲ)-AA on 8 physiological indices of three crops, Zea mays L., Oryza sativa L. and Triticum aestivum L. were studied under simulated acid rain pollution. The results showed that pre-application of La(Ⅲ)-AA could effectively reduce the plasma membrane permeability (45.95%—54.22%), ROS production rate (28.44%—42.31%), malondialdehyde content (31.03%-38.71%), proline content (36.23%—47.17%) and peroxidase activity (48.3%—57.92%). Meanwhile, catalase activity (29.43%—78.61%), chlorophyll content (7.89%—18.76%) and nitrogen metabolism intensity (21.37%—60.63%) were increased. In conclusion, La(Ⅲ)-AA exhibits protective effect against the peroxidative stress caused by acid rain, and pre-spraying La(Ⅲ)-AA can reduce the damage of acid rain pollution to crops.
-
Key words:
- La(Ⅲ)-AA /
- simulated acid rain /
- crops /
- physiological indicators.
-

-
表 1 实验组叶片喷施方式
Table 1. Spraying method of leaves before experiment
时间
Period对照组
BK Group酸雨组
AR Group叶喷组
BS Group第一天 蒸馏水 蒸馏水 20 mg·L−1 La(Ⅲ)-AA 第二天 蒸馏水 酸雨(pH 3.0) 酸雨(pH 3.0)  下载: 导出CSV
下载: 导出CSV
-
[1] 张秀玲, 胡冉, 孙颖, 等. 外源钙对酸雨胁迫下花生幼苗生理特性的影响 [J]. 花生学报, 2019, 48(4): 54-57,62. doi: 10.14001/j.issn.1002-4093.2019.04.009 ZHANG X L, HU R, SUN Y, et al. Effects of calcium on the physiological characteristics of Arachis hypogaea seedlings under the stress of acid rain [J]. Journal of Peanut Science, 2019, 48(4): 54-57,62(in Chinese). doi: 10.14001/j.issn.1002-4093.2019.04.009
[2] 冯宗炜. 中国酸雨的生态影响和防治对策[J]. 云南环境科学, 2000, 19(S1): 1-6. FENG Z W. Ecological Effects and control strategies of acid deposition on ecosystems in China[J]. Yunnan Environmental Science, 2000, 19(Sup 1): 1-6(in Chinese).
[3] 张轩波, 王丽红, 周青. 酸雨胁迫下镧在辣根累积及其对细胞结构的影响 [J]. 稀土, 2017, 38(1): 1-6. doi: 10.16533/J.CNKI.15-1099/TF.201701001 ZHANG X B, WANG L H, ZHOU Q. Accumulation of lanthanum in horseradish roots under acid rain stress and effect on cellular structure [J]. Chinese Rare Earths, 2017, 38(1): 1-6(in Chinese). doi: 10.16533/J.CNKI.15-1099/TF.201701001
[4] 陈俊彤, 谢恩耀, 金琎. 五酸钾对酸雨胁迫下茼蒿的防护 [J]. 环境化学, 2020, 39(10): 2666-2672. doi: 10.7524/j.issn.0254-6108.2019061704 CHEN J T, XIE E Y, JIN J. Potassium pyridine-2, 3, 4, 5, 6-pentacarboxylate protects Artemisia selengensis under acid rain stress [J]. Environmental Chemistry, 2020, 39(10): 2666-2672(in Chinese). doi: 10.7524/j.issn.0254-6108.2019061704
[5] 王锦文, 边才苗. 镧浸种对辣椒种子抗酸雨胁迫能力的强化作用 [J]. 中国稀土学报, 2012, 30(3): 373-379. WANG J W, BIAN C M. Strengthening effects of LaCl3 treatment on resistance of Capsicum seeds to acid rain stress [J]. Journal of the Chinese Society of Rare Earths, 2012, 30(3): 373-379(in Chinese).
[6] 林植芳, 彭长连, 徐信兰, 等. 两种浮萍植物的叶绿体超微结构对模拟酸雨的敏感性 [J]. 热带亚热带植物学报, 2005, 13(3): 217-223. doi: 10.3969/j.issn.1005-3395.2005.03.005 LIN Z F, PENG C L, XU X L, et al. The sensitivity of chloroplast ultrastructure of two duckweed species to simulated acid rain [J]. Journal of Tropical and Subtropical Botany, 2005, 13(3): 217-223(in Chinese). doi: 10.3969/j.issn.1005-3395.2005.03.005
[7] 谢恩耀, 顾嘉豪, 饶福清, 等. 新型化合物对六种植物种子萌发及生理指标的影响 [J]. 分子植物育种, 2019, 17(5): 1703-1710. doi: 10.13271/j.mpb.017.001703 XIE E Y, GU J H, RAO F Q, et al. Effects of new compounds on seed germination and physiological indicator of six plants [J]. Molecular Plant Breeding, 2019, 17(5): 1703-1710(in Chinese). doi: 10.13271/j.mpb.017.001703
[8] ZHANG C, YI X, ZHOU F, et al. Comprehensive transcriptome profiling of tea leaves (Camellia sinensis) in response to simulated acid rain [J]. Scientia Horticulturae, 2020, 272: 109491. doi: 10.1016/j.scienta.2020.109491 [9] 丁士明, 张自立, 梁涛, 等. 外源稀土对土壤中稀土和重金属可交换态的影响 [J]. 环境科学, 2003, 24(4): 122-126. doi: 10.3321/j.issn:0250-3301.2003.04.024 DING S M, ZHANG Z L, LIANG T, et al. Effect of extraneous rare earths (REs) on form of soil exchangeable REs and heavy metals [J]. Chinese Journal of Environmental Science, 2003, 24(4): 122-126(in Chinese). doi: 10.3321/j.issn:0250-3301.2003.04.024
[10] 王蕊. 镧、铈对铜胁迫下豌豆种子萌发和幼苗生长的影响[D]. 雅安: 四川农业大学, 2013. WANG R. Effect of lanthanum, cerium on peas seeds germination and seedling growth under copper stress[D]. Yaan: Sichuan Agricultural University, 2013(in Chinese).
[11] 金琎, 周静. 铈、镧对酸雨胁迫下玉米保护酶影响的比较 [J]. 江苏农业科学, 2012, 40(1): 74-76. doi: 10.3969/j.issn.1002-1302.2012.01.025 JIN J, ZHOU J. Cerium, anthanum comparing corn protective enzyme under acid rain stress effect [J]. Jiangsu Agricultural Sciences, 2012, 40(1): 74-76(in Chinese). doi: 10.3969/j.issn.1002-1302.2012.01.025
[12] 李月福. 轻稀土镧和重稀土钇对玉米和油菜幼苗生长影响研究[D]. 赣州: 江西理工大学, 2013. LI Y F. Effects of light rare earth lanthanum and heavy rare earth yttrium on maize and rape seedling growth [D]. Ganzhou: Jiangxi University of Science and Technology, 2013(in Chinese).
[13] 徐义英, 金琎. 稀土微肥对酸雨胁迫下水稻幼苗期Ⅰ的影响 [J]. 基因组学与应用生物学, 2017, 36(9): 3893-3900. doi: 10.13417/j.gab.036.003893 XU Y Y, JIN J. Effects of rare earth fertilizer on rice seedling stage Ⅰ under acid rain stress [J]. Genomics and Applied Biology, 2017, 36(9): 3893-3900(in Chinese). doi: 10.13417/j.gab.036.003893
[14] 唐加红, 杨玉兰, 苑中原, 等. 镧对干旱胁迫下小麦幼苗抗氧化系统的影响 [J]. 稀土, 2011, 32(1): 12-16. doi: 10.3969/j.issn.1004-0277.2011.01.003 TANG J H, YANG Y L, YUAN Z Y, et al. Effect of lanthanum on antioxidative system in wheet seedlings under drought stress [J]. Chinese Rare Earths, 2011, 32(1): 12-16(in Chinese). doi: 10.3969/j.issn.1004-0277.2011.01.003
[15] 张杰, 黄永杰, 周守标. 铜胁迫下镧对水稻幼苗生长及抗氧化酶活性的影响 [J]. 环境化学, 2010, 29(5): 932-937. ZHANG J, HUANG Y J, ZHOU S B. Effect of La3+ on growth and activities of antioxidant enzymes in rice seedlings under copper stress [J]. Environmental Chemistry, 2010, 29(5): 932-937(in Chinese).
[16] 巩东辉, 王志忠, 季祥. 重金属铅胁迫下镧对螺旋藻生长及生理特性的影响 [J]. 中国稀土学报, 2015, 33(4): 487-493. GONG D H, WANG Z Z, JI X. Effects of lanthanum on growth and physiological characteristics of Spirulina under lead stress [J]. Journal of the Chinese Society of Rare Earths, 2015, 33(4): 487-493(in Chinese).
[17] SATTI Z, AKHTAR M, MAZHAR N, et al. Changes in selected elements of soils under simulated acid rain conditions [J]. Biointerface Research in Applied Chemistry, 2020, 11(4): 11737-11746. doi: 10.33263/BRIAC114.1173711746 [18] 张万萍, 须海丽, 张燕, 等. 喷施稀土对黄瓜幼苗根系活力和矿质元素吸收的影响 [J]. 山地农业生物学报, 2003, 22(6): 512-515. doi: 10.3969/j.issn.1008-0457.2003.06.009 ZHANG W P, XU H L, ZHANG Y, et al. Effects of spraying Rare-earth elements on root activity and main mineral elements of cucumber seedlings [J]. Journal of Mountain Agriculture and Biology, 2003, 22(6): 512-515(in Chinese). doi: 10.3969/j.issn.1008-0457.2003.06.009
[19] 余海兵, 刘正, 王波. 农用稀土对糯玉米幼苗光合变化和生理指标的分析 [J]. 中国稀土学报, 2011, 29(1): 119-124. YU H B, LIU Z, WANG B. Analysis of rare earths in agriculture on photosynthetic changes and physiological indicators in waxy corn seedings [J]. Journal of the Chinese Rare Earth Society, 2011, 29(1): 119-124(in Chinese).
[20] 贾金涛, 余悦旗, 武孟云, 等. 氯化镧对玉米幼苗抗盐性的影响 [J]. 西北农业学报, 2015, 24(5): 64-68. doi: 10.7606/j.issn.1004-1389.2015.05.011 JIA J T, YU Y Q, WU M Y, et al. Effect of lanthanum chloride on salt tolerance of maize seedlings [J]. Acta Agriculturae Boreali-Occidentalis Sinica, 2015, 24(5): 64-68(in Chinese). doi: 10.7606/j.issn.1004-1389.2015.05.011
[21] 张利霞, 常青山, 侯小改, 等. 盐胁迫下镧对苏丹草种子发芽的影响 [J]. 种子, 2014, 33(7): 78-81. doi: 10.3969/j.issn.1001-4705.2014.07.021 ZHANG L X, CHANG Q S, HOU X G, et al. Effects of La3+ on germination characteristics of the Sorghum sudonense seed under salt stress [J]. Seed, 2014, 33(7): 78-81(in Chinese). doi: 10.3969/j.issn.1001-4705.2014.07.021
[22] 任鹏辉. 叶面喷施硝酸镧(La(NO3)3)对NaCl胁迫下紫花苜蓿(Medicago sativa)生理活性的影响[D]. 杨凌: 西北农林科技大学, 2017. REN P H. Effect of La(NO3)3 on physiological activity in alfalfa(Medicago sativa)under NaCl stress[D]. Yangling: Northwest A & F University, 2017(in Chinese).
[23] 张梅, 刘曙光, 史向群, 等. 氨基酸及氨基酸螯合肥生物学功效研究进展[J]. 湖北农业科学, 2019, 58(S1): 1-3, 6. ZHANG M, LIU S G, SHI X Q, et al. Biological effect of plant amion acids and amino acids trace-element fertilizers[J]. Hubei Agricultural Sciences, 2019, 58(Sup 1): 1-3, 6(in Chinese).
[24] 庄钟娟, 肖艳, 揣俊峰, 等. 冲施氨基酸液肥对小油菜生长及养分累积的影响 [J]. 中国园艺文摘, 2016, 32(12): 53,192. doi: 10.3969/j.issn.1672-0873.2016.12.018 ZHUANG Z J, XIAO Y, CHUAI J F, et al. Effects of liquid amino acid fertilizer on growth and nutrient accumulation of rapeseed [J]. Chinese Horticulture Abstracts, 2016, 32(12): 53,192(in Chinese). doi: 10.3969/j.issn.1672-0873.2016.12.018
[25] 胡相, 程小龙, 袁国明. 氨基酸叶面肥和农抗9510在烟草上的应用 [J]. 植物医生, 2014, 27(4): 42-44. doi: 10.13718/j.cnki.zwys.2014.04.023 HU X, CHENG X L, YUAN G M. Application of amino acid foliar fertilizer and agricultural resistance 9510 in tobacco [J]. Plant Doctor, 2014, 27(4): 42-44(in Chinese). doi: 10.13718/j.cnki.zwys.2014.04.023
[26] 曹焱. 氨基酸水溶肥对草莓生理特性的影响 [J]. 农业科技与信息, 2021(16): 42-44. doi: 10.3969/j.issn.1003-6997.2021.16.017 CAO Y. Effects of amino acid water-soluble fertilizer on physiological characteristics of strawberry [J]. Agricultural Science-Technology and Information, 2021(16): 42-44(in Chinese). doi: 10.3969/j.issn.1003-6997.2021.16.017
[27] 郭丽颖, 赵宏伟, 王敬国, 等. 黑龙江省稻瘟病菌生理小种鉴定和主栽水稻品种抗病性及遗传多样性分析 [J]. 核农学报, 2015, 29(8): 1444-1454. doi: 10.11869/j.issn.100-8551.2015.08.1444 GUO L Y, ZHAO H W, WANG J G, et al. Identification of physiological race of rice blast fungus and disease resistance and genetic diversity analysis on major cultivars in Heilongjiang Province [J]. Journal of Nuclear Agricultural Sciences, 2015, 29(8): 1444-1454(in Chinese). doi: 10.11869/j.issn.100-8551.2015.08.1444
[28] 许猛. 复合氨基酸制剂对小白菜和棉花抗逆性的影响[D]. 北京: 中国农业科学院, 2018. XU M. Effects of a compound amino acid preparation on stress resistance of pak choi and cotton[D]. Beijing: Chinese Academy of Agricultural Sciences, 2018(in Chinese).
[29] ZHONG Y Q, CHEN J J. Ameliorative effects of Lanthanum(Ⅲ) on Copper(Ⅱ) stressed rice (Oryza sativa) and its molecular mechanism revealed by transcriptome profiling [J]. Plant Physiology and Biochemistry, 2020, 152: 184-193. doi: 10.1016/j.plaphy.2020.05.004 [30] 周芸. 花生施用氨基酸稀土微肥效果试验 [J]. 农业研究与应用, 2013(3): 20-22. doi: 10.3969/j.issn.2095-0764.2013.03.006 ZHOU Y. Effect of rare earth amino acid microfertilizer on peanut [J]. Agricultural Research and Application, 2013(3): 20-22(in Chinese). doi: 10.3969/j.issn.2095-0764.2013.03.006
[31] 罗子渝, 任维莉. 酸胁迫对冬青叶片叶绿素含量和膜透性的影响 [J]. 农业与技术, 2020, 40(3): 25-27. doi: 10.19754/j.nyyjs.20200215009 LUO Z Y, REN W L. Effects of acid stress on chlorophyll content and membrane permeability of Holly leaves [J]. Agriculture and Technology, 2020, 40(3): 25-27(in Chinese). doi: 10.19754/j.nyyjs.20200215009
[32] 李小方, 张志良. 植物生理学实验指导[M]. 5版. 北京: 高等教育出版社, 2016: 218-227. LI X F, ZHANG Z L. Laboratory physiology experiment guide [M]. (fifth edition)Beijing: Higher Education Press, 2016: 218-227(in Chinese).
[33] 阮方毅. 稀土氨基酸类配合物的制备及应用研究[D]. 西安: 陕西科技大学, 2019. RUAN F Y. Preparation and application of rare earth amino acids complexes[D]. Xi'an: Shaanxi University of Science & Technology, 2019(in Chinese).
[34] 姚梦婕, 金琎. 铈对酸雨胁迫下水稻幼苗期防护与修复作用 [J]. 环境化学, 2016, 35(12): 2553-2558. doi: 10.7524/j.issn.0254-6108.2016.12.2016051804 YAO M J, JIN J. Effects of cerium on the protection and repair of rice seedlings under acid rain stress [J]. Environmental Chemistry, 2016, 35(12): 2553-2558(in Chinese). doi: 10.7524/j.issn.0254-6108.2016.12.2016051804
[35] 侯明, 张兴龙, 路畅, 等. V(Ⅴ)、Cr(Ⅵ)单一和复合胁迫对小麦幼苗生长和生理特性的影响 [J]. 环境化学, 2012, 31(7): 1016-1022. HOU M, ZHANG X L, LU C, et al. Effects of of V, Cr single and combined stress on the growth and physiological characteristics of wheat seedling [J]. Environmental Chemistry, 2012, 31(7): 1016-1022(in Chinese).
[36] ZHAI J L, XU H X, CONG X L, et al. Ca2+/H^+ exchange in the plasma membrane of Arabidopsis thaliana leaves [J]. Acta Physiologiae Plantarum, 2013, 35(1): 161-173. doi: 10.1007/s11738-012-1059-y [37] 孙静雯, 王丽红, 周青. 酸雨对水稻不同生育期叶片叶绿体ATP酶的影响 [J]. 安全与环境学报, 2017, 17(5): 1895-1899. doi: 10.13637/j.issn.1009-6094.2017.05.053 SUN J W, WANG L H, ZHOU Q. Effect of acid rain on chloroplast ATP enzyme in rice at different growth stages [J]. Journal of Safety and Environment, 2017, 17(5): 1895-1899(in Chinese). doi: 10.13637/j.issn.1009-6094.2017.05.053
[38] 王丽红, 周青, 曾庆玲. 3类抗性种子萌发过程中糖代谢对酸雨胁迫的响应 [J]. 环境科学, 2008, 29(3): 799-803. doi: 10.3321/j.issn:0250-3301.2008.03.043 WANG L H, ZHOU Q, ZENG Q L. Reponses of sugar metabolism in seed germination of three various acid-fast plants to acid rain [J]. Environmental Science, 2008, 29(3): 799-803(in Chinese). doi: 10.3321/j.issn:0250-3301.2008.03.043
-
 点击查看大图
点击查看大图

计量
- 文章访问数: 1857
- HTML全文浏览数: 1857
- PDF下载数: 73
- 施引文献: 0


