

-
水华(algal bloom)是在一定的营养、气候和水文条件下藻类等浮游生物大量繁殖并在水体表面聚集,使水体颜色发生变化的现象[1]. 水华的发生可能引起严重的生态环境问题,大量繁殖的藻类死亡后耗氧分解会造成水体缺氧,这严重威胁了水体中其他生物的生存,从而导致生态失衡,而如果饮用水源发生水华则会影响饮用水安全,造成严重的经济损失[2]. 一直以来,水华都是国内外研究学者重点关注的生态环境问题之一.
湖光岩玛珥湖(以下简称湖光岩)是距今14—16万年前由平地火山喷发后火山口下沉形成的湖泊,是世界上最大且保存最完整的玛珥湖[3]. 湖光岩是封闭性湖泊,四周被火山碎屑岩包围,且没有河流的注入与流出,湖水水位的变化主要取决于大气降水和地下水位的变化[4]. 前人有关湖光岩生态问题的研究主要集中在水体营养盐的时空分布[3]、浮游植物对溶解态氮的吸收[5]以及浮游植物种属的季节性变化[6]等方面,而有关湖光岩水华问题的研究尚未有过报道. 早在2009年以及2011年便有新闻报道关于湖光岩的湖面漂浮着蓝藻,但当时却未引起各学界的重视[7]. 2021年12月—2022年3月,笔者注意到湖光岩的湖面漂浮着大量的浮游藻类,东、北部水域的湖水浑浊且湖滩被一层绿色的藻泥所覆盖,此外在东、北部湖滩也发现了不少已经死亡的乌龟(图1),经观察确认湖光岩暴发了水华. 鉴于有关湖光岩水华现象的研究尚未有过报道,故本文对湖光岩的水华现象开展初步的研究分析,以期为认识湖光岩水华发生的机理提供科学依据.
湖光岩为封闭性湖泊,是研究认识较为单纯的水文条件下水华形成与发展的理想场所,对研究封闭性湖泊水华的发生具有独特的科学意义. 此外,湖光岩为国家4A级旅游景点,水华的发生必然对湖泊水体及周围的生态环境有所影响,因此分析和探讨湖光岩发生水华的原因具有积极的理论与实际意义. 本研究通过对湖光岩水华暴发的主要藻种鉴定以及水体样品的水质分析等,试图从营养盐、水文条件以及气象因素等方面探讨分析湖光岩发生水华的原因,以期认识玛珥湖水华发生的影响控制因素,同时也为湖光岩水华的治理与预防提供科学依据.
-
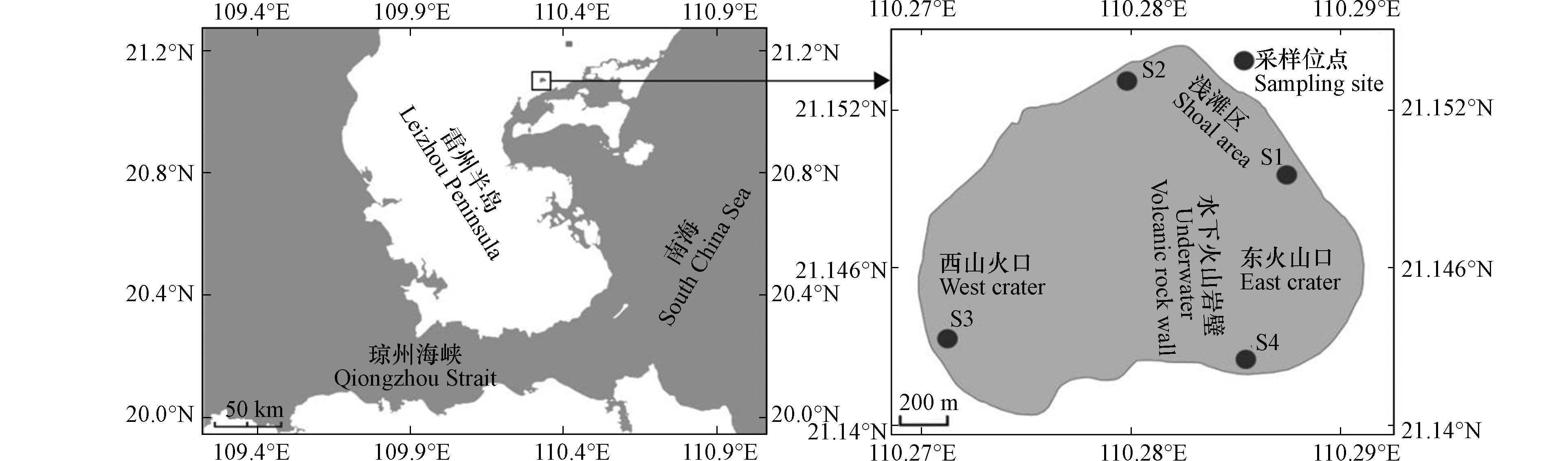
湖光岩(21°09′N,110°17′E)位于中国广东省湛江市,其外形呈心形形状,由紧邻的两个火山喷发口组合而成,在水下被火山岩壁分割成东、西两湖,东湖小且浅,西湖较大且深,湖泊水位较低时可看见东西两湖之间不连续的火山石出落于水中. 湖泊总面积为 2.3 km2,水深最深处约为 22 m[8]. 水华藻类样品及水样的采集主要是结合湖光岩的地形环境以及水华发生的位置进行采样点的布设,如图2所示,其中S1、S2站点位于浅滩区域,水深约1—2 m,该区域发生水华现象,湖水中漂浮着大量的浮游藻类并且湖滩有藻泥沉积覆盖,在S1、S2站点采集水华藻类样品和水样,S3站点位于西湖区域,S4站点位于东湖区域,由于S3、S4站点的区域没有发生水华,故在这两个样点只采集了水样.
-
样品的采集时间为2022年3月,使用采水器取水面以下约 0.5 m深处的水样1 L,完成后立即带回实验室,使用0.45 μm的醋酸纤维滤膜过滤,过滤后的水样冷冻保存于-20 ℃的冰箱,用于开展营养盐(硝态氮(NO3−-N)、亚硝态氮(NO2−-N)、铵态氮(NH4+-N)、磷酸盐(PO43−-P)和硅酸盐(SiO32−-Si))的分析. 另采集250 mL水样,加入3—5 mL的鲁哥试剂固定液,带回实验室经沉淀浓缩后进行镜下观察拍照,用于水华主要藻种的分析与鉴定.
-
湖水理化性质的测定包括现场水体的水温、pH值、盐度的测定,均使用便捷式多参数分析仪(上海雷磁有限公司,DZB—718L)进行现场测定. 营养盐的测定根据《水和废水监测分析方法》[9]进行测定,其中氨氮采用纳氏试剂分光光度法,亚硝酸盐采用萘乙二胺分光光度法,硝酸盐采用紫外分光光度法,磷酸盐采用磷钼蓝分光光度法,硅酸盐采用硅钼黄法分光光度法. 水华主要藻种的鉴定通过光学显微镜对采集的浮游藻类进行镜下观察和拍照,并参照《中国淡水藻类》[10]、《淡水微型生物图谱》[11]进行鉴定.
-
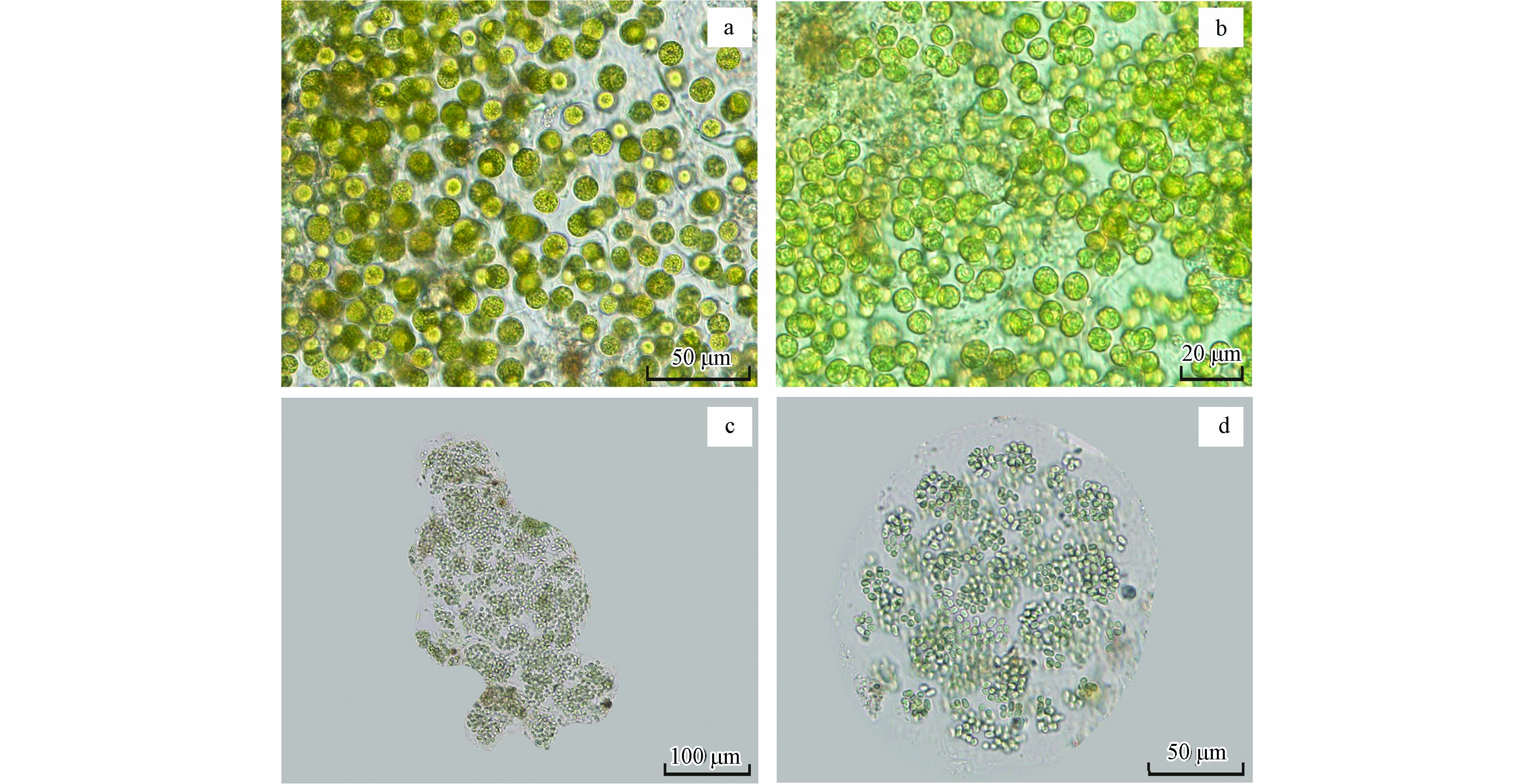
显微镜观察显示,构成湖光岩水华的藻种主要有两种,分别显示于图3a、b和图3c、d. 其中图3a、b显示的藻类为球形单细胞体,呈黄绿色,细胞壁薄,细胞直径5—10 μm;参照《中国淡水藻类》[10]和《淡水微型生物图谱》[11]进行鉴定,确定该藻种为绿藻门(Chlorophyta)、绿藻纲(Chlorophyceae)、绿球藻目(Chlorococcales)、小球藻科(Chlorellaceae)、小球藻属(Chlorella)中的小球藻(Chlorella vulgaris). 图c、d显示的藻类为多数细胞组成的群体,成熟的群体明显裂开,胶被的某些区域破裂或穿孔;群体中的细胞呈球形或椭圆形,直径3—7 μm,呈蓝绿色;经图谱鉴定该藻种为蓝藻门(Cyanophyceae)、蓝藻纲(Cyanophyceae)、色球藻目(Chroococcales)、色球藻科(Chroococcaceae)、微囊藻属(Microcystis)中的铜绿微囊藻(Microcystis aeruginosa)[12]. 因此,引起湖光岩水华暴发的主要藻类为绿藻和蓝藻,且以绿藻占据优势,通过显微镜观察小球藻的数量占比可达60%.
张才学等[6]于2006年对湖光岩的浮游藻类进行周年的调查,结果检出湖光岩水体中浮游藻类260种,以绿藻门、蓝藻门、硅藻门为主,其中绿藻门占45%,硅藻门占30.4%,蓝藻门占16.5%,绿藻门的小球藻以及蓝藻门的小型色球藻、水华微囊藻、煤黑厚皮藻为全年广布优势种;2013年张国维等[13]检出湖光岩水体中浮游藻类135种,其中绿藻门占33.3%,蓝藻门占30.4%,硅藻门占22.2%,绿藻门的小球藻、蓝藻门的水华微囊藻、铜绿微囊藻以及硅藻门的颗粒直链藻为全年广布优势种. 可见,由于环境理化因子以及水质营养条件的改变,水华期间与没有发生水华期间浮游藻类的组成结构发生改变.
-
表1为2022年3月份湖光岩各站点理化状态的测定结果. 由表1可看出,湖光岩各站点的温度变化范围为25.10—29.20 ℃,平均温度为26.72 ℃,且水华发生区域的S1、S2站点的水温要比S3、S4站点的水温高2—4 ℃,这与湖光岩的地理环境有关,S3、S4站点位于湖光岩的南部,南部的植被较高且密集分布,阳光易被遮挡,因此湖泊的南部日照时间较短,接收的阳光较少,而S1、S2站点位于湖光岩的东北部,在一天中能接收更充足的阳光,因而S1、S2站点的水温更高[14].
温度是水体环境中最为重要的参数,也是诱发水华暴发的重要因素之一. 温度通过影响藻类的代谢强度和光合作用,控制着藻类的生长、发育和分布等[15]. 此次构成湖光岩水华暴发的两种主要藻种中,小球藻对温度的适应范围较广,而铜绿微囊藻在28—32 ℃的水温条件下生长速率最高,更适宜在较高温的条件下生长[16-17]. 由于湖光岩水华的暴发主要发生于冬春季节,冬季的湖水温度相对偏低,更适合小球藻的生长繁殖,这应是湖光岩水华暴发的主要藻种中小球藻占据优势的主要原因. 而发生水华区域的S1、S2站点位于湖光岩的北部,冬季期间能接收更充足的阳光,水温比位于日照少的南部区域的S3、S4站点要高,更有利于藻类进行光合作用.
-
表1显示湖光岩各站点的pH值变化范围为7.98—8.67,水质偏弱碱性,其中S2站点的pH值最高为8.67,而且S1、S2站点的pH值要比S3、S4站点的pH值高,这与浮游藻类在生长过程中吸收水体中的CO2进行光合作用有关,使得水体中的氢离子浓度降低,pH值升高. 一般情况下,藻类适宜生长在中性或者弱碱性的环境中,而水体的pH值也会随着藻类的增多而升高[18]. 小球藻对pH值的适应范围较广,适应能力较强. 而铜绿微囊藻在pH值为8.0—9.5的环境条件下生长繁殖能力较强,pH值过低或过高都会对铜绿微囊藻的生长有所影响[19]. 总体上看,湖光岩的酸碱度条件有利于小球藻和铜绿微囊藻的生长繁殖.
-
由表1可见,湖光岩各站点不同营养盐的含量中,SiO32−-Si的平均含量最高。4个站点的SiO32−-Si含量变化较大,范围为0.141—4.140 mg·L−1,S1站点的SiO32−-Si含量最高,为4.140 mg·L−1. PO43−-P的含量最低,含量范围为0.001—0.004 mg·L−1,其中S1站点的PO43−-P含量最高为0.004 mg·L−1. 在4个站点中NO2−-N的含量最稳定,4个站点的NO2−-N含量都为0.003 mg·L−1. NO3−-N的含量在4个站点中基本稳定,其中S1站点的NO3−-N含量最高,为0.192 mg·L−1. NH4+-N的含量变化在4个站点变化不大,变化范围为0.097—0.137 mg·L−1,其中S1站点的NH4+-N含量最高,为0.137 mg·L−1.
不同时期下湖光岩表层水营养盐含量的对比分析表明(表2),SiO32−-Si的含量在不同时期测定的营养盐组分中都是最高的,变化的范围为0.2674—1.5360 mg·L−1,PO43−-P含量最低,变化范围为0.0006—0.0030 mg·L−1. 图4为不同时期湖光岩表层水营养盐含量以及本研究不同采样点营养盐含量柱形图. 由图4中可明显看出,除了NO3−-N之外,本研究测定的NH4+-N、NO2−-N、PO43−-P、SiO32−-Si含量远高于前人测定的数值. 此外,从图4中可看出,除NO2−-N之外,水华发生区域S1、S2站点测定的NH4+-N、NO3−-N、PO43−-P、SiO32−-Si含量均高于S3、S4站点的含量. 营养盐是影响水体中藻类生长发育的重要条件,并在一定程度上导致水体中的优势藻类群落发生改变. 由此可见,相对较高的营养盐含量是引起湖光岩水华暴发的主要原因.
在分析的5种营养盐中,SiO32−-Si的含量最高,一方面这可能是由于湖泊底层沉积物中的有机质氧化分解产生的,另一方面可能是因为湖光岩四周为火山碎屑岩,火山碎屑岩的硅元素含量较高,岩石中的可溶性物质经雨水冲刷被带入水体中,从而使水体中的SiO32−-Si含量背景值较高[3]. 已有研究发现在浮游藻类中蓝藻和绿藻对磷有较高的需求,对硅没有明显的需求,而硅藻则对硅的需求较高[21-22]. 引发湖光岩水华暴发的两种主要藻类属于蓝藻和绿藻,因此,较高含量的SiO32−-Si对水华的形成没有明显的促进作用,其含量也可能没有达到让湖光岩水体暴发硅藻水华的浓度门限.
在本研究分析的5种营养盐中,PO43−-P含量最低,贝格最小值定律指出,植物生长取决于外界提供给它的所需养料中数量最少的一种[2]. Redfield[23]指出浮游藻类吸收利用氮、磷营养盐的最适宜比例为16:1,在研究中一般认为水体中的氮磷比大于16:1存在磷限制,而小于16:1则存在氮限制. 由表1可见,本研究中4个站点的N:P变化范围为83—227(N = DIN,P = PO43−-P),其中S3站点的氮磷比值最高为227,S1站点的氮磷比值最低为83,表现出明显的磷限制,这一分析结果与前人的研究结果一致[3, 6]. 且由表2及图4可见,相比于前人的研究,本研究的氮磷比相对较低. 因此,相比于氮营养盐含量的增加,磷营养盐含量的增加更有利于浮游藻类的生长繁殖. 由图4可见,相比于前人的研究,本研究的PO43−-P含量相对偏高,且水华发生区域的S1、S2站点的PO43−-P含量要比S3、S4站点的PO43−-P含量高. 因此,PO43−-P含量增加是导致湖光岩发生水华的重要因子之一.
张国维等[5]利用15N 稳定同位素示踪技术研究湖光岩浮游藻类对氮的吸收速率,结果发现湖光岩的浮游藻类对NH4+-N的吸收速率最高,更偏好利用NH4+-N,而对NO3−-N、NO2−-N则具有一定的亲和力. 由此推测,NH4+-N 也是调控湖光岩浮游藻类生长的重要营养盐因子之一. 由图4可知,相比于之前没有发生水华现象的研究,本研究中NH4+-N的含量相对较高,而且相比于S3、S4站点,水华发生区域的S1、S2站点的NH4+-N含量也是相对偏高的. 根据以上分析可见,水体中PO43−-P、NH4+-N含量的增加是造成水华现象发生的主要原因.
湖光岩为封闭性湖泊,其营养物质的循环主要是生态系统的内部循环,而外源营养物质的输入较少[6]. 湖光岩的营养物质内部循环受水体温度变化的影响较大,湖光岩的水体温度在垂向上具有季节性分层的现象,夏季表层水的温度高,表层水与较冷的深层水出现分层,阻碍了表层水与深层水及底层沉积物的营养盐循环交换,而到了冬春季节表层水的温度开始下降,水体的分层被破坏,水体发生混合,表层水的营养盐会有所增加[3, 24]. 此次湖光岩水华的暴发正值冬春季节,推测PO43−-P、NH4+-N等营养盐含量的增加主要是由于表层水与深层水发生垂直混合,从而使表层水的营养盐含量增加. 且S1、S2站点位于湖光岩浅滩区域,水深较浅,在冬季风的作用下其水体垂直混合更充分,因而其营养盐含量比S3、S4站点的高,更有利于水华的暴发.
-
浮游藻类的生长不仅受温度、光照、pH、营养盐、微量元素等影响因子的控制,水位的变化对藻类的生长也具有重要的调节作用[25]. 水位下降所引起的垂向轻微扰动能够使水体中的营养盐获得重新分配,激发底层水体和沉积物中营养盐的活性,为浮游藻类的生长提供了更持久的营养盐支撑条件[26]. 此外,水位的下降增强了水体的透光性,有利于浮游藻类进行光合作用,为浮游藻类的生长提供了有利的条件.
根据湛江市水文局的水文记录显示,在没有发生水华的年份湖光岩的平均水位多保持在20—21 m,而2021年以来湖光岩的水位在持续走低,2021—2022年最低水位出现在2022年4月中旬,为18.93 m,这表明在水华发生期间湖光岩的水位下降了近2 m. 湖光岩为封闭性湖泊,没有河流的注入与流出,水位的变化主要是取决于降雨和地下水位的变化. 湖光岩的气候属于典型的热带季风气候,干湿季分明,降雨主要集中在夏秋季节,冬春季节降雨量少,偏干旱[3]. 可见在冬春季节,水华发生期间降雨量偏少,减少了湖水的主要来源,长时间的干旱加快了湖水的蒸发速率,造成水位降低. 水位的下降使得水体中的营养盐得以重新分配,增强水体的透光性,为藻类的生长创造有利的条件. 此外,湖光岩为封闭性湖泊,在水位下降的条件下,水体中的营养盐得以富集,且更有利于水体的垂直混合,使表层的营养盐含量增加,加剧水华的暴发进程.
-
(1)引发2021—2022年湖光岩水华暴发的主要藻种为绿藻门中的小球藻以及蓝藻门中的铜绿微囊藻,其中小球藻的数量占比可达60%.
(2)2021—2022年湖光岩水华暴发的主要原因是在冬春季节,表层水体的温度下降,水体的分层被破坏,表层水体与深层水及底层沉积物的营养盐发生内部循环,导致表层水营养盐浓度增高,特别是PO43−-P和NH4+-N的增加,再加上适宜的水温、pH、光照条件以及水位的下降,为藻类的生长创造有利的条件,造成水华的发生. 因此,2021—2022年湖光岩水华的暴发是营养盐、水体环境因子以及气候因子等影响因子综合作用的结果.
致谢:感谢广东省湛江市水文局提供湖光岩水位数据资料和罗涛在藻类样品采集中给予的帮助,特别感谢广东海洋大学张才学教授和南宁师范大学徐轶肖教授对论文提出的宝贵修改意见,在此向他们表示衷心的感谢!
引发2021—2022年度湖光岩玛珥湖水华事件的主要藻种及原因分析
The dominant algal species and the controlling factors triggered 2021—2022 Huguangyan Maar Lake algal bloom event
-
摘要: 水华(algal bloom)是水体中藻类过度繁殖所引起的生态环境问题,水华的暴发会对水体环境以及水生生态平衡造成严重的破坏. 湖光岩玛珥湖是低平地区火山口形成的湖泊,具有相对封闭的水体环境,是研究认识较为单纯的水文条件下水华形成与发展的理想场所,研究其水华暴发期间的主要藻种构成及水生态环境条件对认识水华形成的原因具有积极的意义. 本文对2021年12月—2022年3月期间湖光岩水华暴发的主要藻类进行了分析鉴定,并从营养盐条件、水文条件、气象因素等方面探讨分析湖光岩水华暴发的原因. 结果表明:(1)湖光岩水华暴发的主要藻种为绿藻门中的小球藻(Chlorella vulgaris)以及蓝藻门中的铜绿微囊藻(Microcystis aeruginosa);(2)湖光岩表层水营养盐含量分析显示,SiO32−-Si的含量最高,PO43−-P的含量最低,NO2−-N、NO3−-N、NH4+-N的含量在不同采样点基本保持稳定,而与前人在没有发生水华的年份所做的湖光岩水体营养盐含量分析相比,水华期间SiO32−-Si、PO43−-P以及NH4+-N的含量显著增加,而其他营养盐组分则变化不大,PO43−-P、NH4+-N为调控湖光岩浮游藻类生长的重要营养盐因子,因此水体中PO43−-P、NH4+-N含量的增加是造成湖光岩发生水华的主要原因;(3)湖光岩水华的暴发主要与水体中营养盐的浓度变化有关,而适宜的水文环境以及气候条件等也有利于水华的暴发.Abstract: Algal bloom is an ecological and environmental problem caused by the overbreeding of algal in the water, and the outbreak of algal bloom will cause serious damage to the water environment and aquatic ecological balance. Huguangyan Maar lake is a lake formed by volcanic craters in low-flat areas, with a relatively closed hydrological environment, it is an ideal place to study the occurrence of algal bloom under relatively simple hydrological conditions, and the study of the main algal species and ecological conditions of lake water environmental conditions during the outbreaks of algal bloom is significance for understanding the causes of algal bloom. This study has analyzed and identified the dominant algal species which caused the algal bloom in Huguangyan Maar lake from December 2021 to March 2022, and discussed the outbreak causes of algal bloom in terms of nutrient conditions, hydrological conditions, meteorological factors. The results showed that (1) the dominant algal species that consisting of the algal bloom are chlorella vulgaris of the phylum Chlorellavulgaris and microcystis aeruginosa of the cyanobacteria phylum; (2) the analysis of the surface water nutrients show that the content of SiO32−-Si is the highest nutrient as the PO43−-P being the lowest one; the content of NO2−-N、NO3−-N、NH4+-N is basically same at all four sampling sites. By comparing with the earlier nutrient data of the surface water in Huguangyan Maar lake in previous studies, the content of SiO32−-Si, PO43−-P and NH4+-N show significant increase during the algal bloom, while other nutrients are generally remain same, PO43−-P、NH4+-N are important nutrient factors regulating the growth of the planktonic algal in Huguangyan Maar lake, therefore it seems that the increase content of PO43−-P and NH4+-N in the lake water are likely the main reason for the outbreak of algal bloom in Huguangyan Maar lake; (3) the outbreak of algal bloom in Huguangyan Maar lake is mainly related to the change of nutrients concentration in water, and the appropriate hydrological environment and climatic conditions aggravate the outbreak process.
-
Key words:
- algal bloom /
- dominant algal species /
- nutrients of surface water /
- Huguangyan Maar Lake.
-

-

图 1 湖光岩东、北部水域湖水浑浊(a)和岸边死亡的乌龟(b)
Figure 1. Turbid water(a)and dead turtles(b)on the east and northt of the Huguangyan
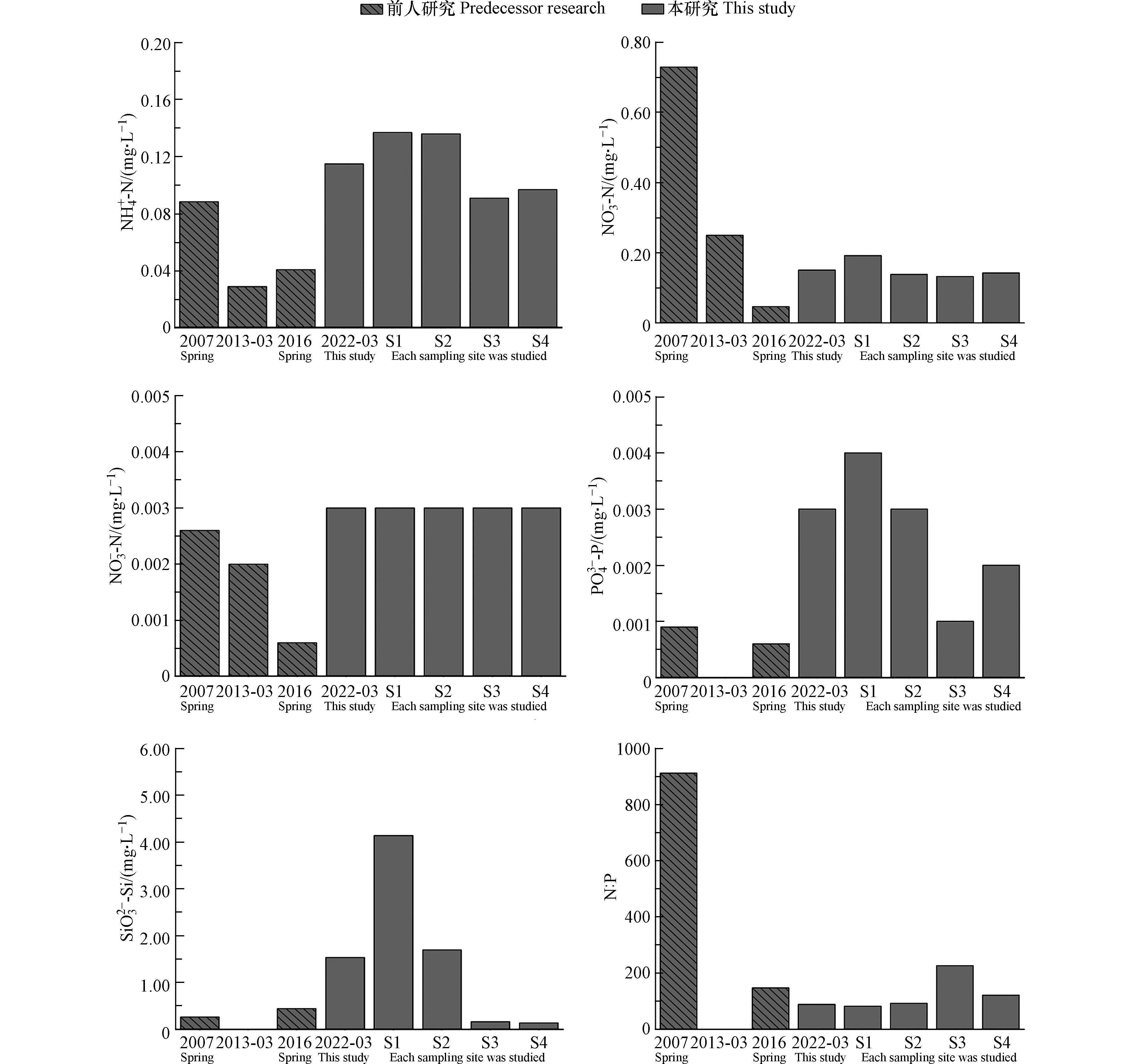
图 4 不同时期湖光岩表层水营养盐含量以及本研究不同采样点营养盐含量柱形图
Figure 4. Column chart of nutrient content in the surface water of the Huguangyan in different periods and nutrient content at different sampling points in this Study
表 1 湖光岩不同采样站点理化因子的测定结果
Table 1. Determination results of physical and chemical factors at various stations of the Huguangyan
站点
StationsT/℃ pH S/% NH4+-N/
(mg·L−1)NO3--N/
(mg·L−1)NO2--N/
(mg·L−1)PO43--P/
(mg·L−1)SiO32--Si/
(mg·L−1)N:P S1 27.4 8.601 0.006 0.137 0.192 0.003 0.004 4.140 83 S2 29.2 8.665 0.006 0.136 0.139 0.003 0.003 1.700 93 S3 25.2 7.980 0.006 0.091 0.133 0.003 0.001 0.164 227 S4 25.1 8.017 0.006 0.097 0.143 0.003 0.002 0.141 122  下载: 导出CSV
下载: 导出CSV
表 2 湖光岩表层水营养盐含量历史对比分析表
Table 2. Table for historical comparison and analysis of nutrient content determination in the surface water of the Huguangyan
时间
TimeT/℃ pH NH4+-N/
(mg·L−1)NO3--N/
(mg·L−1)NO2--N/
(mg·L−1)PO43--P/
(mg·L−1)SiO32--Si/
(mg·L−1)N:P 参考文献
References2007春季 27.00 8.40 0.0885 0.7295 0.0026 0.0009 0.2674 912 [6] 2013.03 20.30 7.88 0.0290 0.2500 0.0020 — — — [20] 2016春季 — 6.85—8.44 0.0410 0.0470 0.0006 0.0006 0.4460 148 [3] 2022.03 26.70 8.32 0.1150 0.1510 0.0030 0.0030 1.5360 89 本研究
下载: 导出CSV
-
[1] 牛晓君. 富营养化发生机理及水华暴发研究进展 [J]. 四川环境, 2006, 25(3): 73-76. NIU X J. Research progress of eutrophication mechanism and breakout of water bloom [J]. Sichuan Environment, 2006, 25(3): 73-76(in Chinese).
[2] 马健荣, 邓建明, 秦伯强, 等. 湖泊蓝藻水华发生机理研究进展 [J]. 生态学报, 2013, 33(10): 3020-3030. doi: 10.5846/stxb201202140200 MA J R, DENG J M, QIN B Q, et al. Progress and prospects on cyanobacteria bloom-forming mechanism in lakes [J]. Acta Ecologica Sinica, 2013, 33(10): 3020-3030(in Chinese). doi: 10.5846/stxb201202140200
[3] 陈法锦, 劳齐斌, 卞培旺, 等. 湖光岩玛珥湖水体中营养盐的时空分布特征及其影响因素 [J]. 湖泊科学, 2018, 30(6): 1693-1706. doi: 10.18307/2018.0621 CHEN F J, LAO Q B, BIAN P W, et al. Spatial and temporal distributions of nutrients and their influencing factors in the Huguangyan Maar Lake [J]. Journal of Lake Sciences, 2018, 30(6): 1693-1706(in Chinese). doi: 10.18307/2018.0621
[4] 柏杨. 湖光岩玛珥湖多指标记录下的全新世气候环境演变[D]. 北京: 中国科学院大学, 2017. BAI Y. A multi-proxy lacustrine record of Holocene climatic and environmental changes in Huguangyan, Maar Lake[D]. Beijing: University of Chinese Academy of Sciences(Guangzhou Institute of Geochemistry, Chinese Academy of Sciences), 2017 (in Chinese).
[5] 张国维, 李长玲, 黄翔鹄, 等. 湖光岩玛珥湖春季浮游植物对溶解态氮的吸收 [J]. 湖泊科学, 2015, 27(3): 527-534. doi: 10.18307/2015.0321 ZHANG G W, LI C L, HUANG X H, et al. Uptake of dissolved nitrogen by phytoplankton in spring in Huguangyan Maar Lake [J]. Journal of Lake Sciences, 2015, 27(3): 527-534(in Chinese). doi: 10.18307/2015.0321
[6] 张才学, 孙省利, 谢少英, 等. 湖光岩玛珥湖的浮游植物 [J]. 水生生物学报, 2008, 32(5): 620-630. ZHANG C X, SUN X L, XIE S Y, et al. The phytoplankton of the Huguangyan maar lake [J]. Acta Hydrobiologica Sinica, 2008, 32(5): 620-630(in Chinese).
[7] 蔡兴争. 湖光岩湖边惊现不明绿物[N]. 湛江日报, 2011-02-14. CAI X Z. Unknown green objects appear at the lakeside of Huguangyan[N]. Zhanjiang Daily, 2011-2-14 (in Chinese).
[8] 王倩, 周浩达, 胡建芳, 等. 湛江湖光岩玛珥湖中长链烷基二醇类化合物的检出及可能的环境意义 [J]. 地球化学, 2013, 42(2): 188-195. WANG Q, ZHOU H D, HU J F, et al. Identification and potential environmental implication of long-chain alkyl diols in Huguangyan maar Lake [J]. Geochimica, 2013, 42(2): 188-195(in Chinese).
[9] 国家环境保护总局《水和废水监测分析方法》编委会. 水和废水监测分析方法 (第四版) [M]. 北京: 中国环境科学出版社, 2002. Editorial Board of Monitoring and Analysis Methods for Water and Wastewater of State Environmental Protection Administration. Monitoring and Analysis Methods for Water and Wastewater (Fourth Edition) [M]. Beijing: China Environmental Science Press, 2002 (in Chinese).
[10] 胡鸿钧, 李尧英, 魏印心, 等. 中国淡水藻类[M]. 上海: 上海科学技术出版社, 1980. HU H J, LI Y Y, WEI Y X, et al. Freshwater algae in China[M]. Shanghai: Shanghai Scientific & Technical Publishers, 1980(in Chinese).
[11] 周凤霞, 陈剑虹. 淡水微型生物图谱[M]. 北京: 化学工业出版社, 2005. ZHOU F X, CHEN J H. Freshwater micro-biological atlas[M]. Beijing: Chemical Industry Press, 2005(in Chinese).
[12] 王然, 朱传雪, 岳俊阳, 等. 巢湖铜绿微囊藻的分离鉴定及其生长特性研究 [J]. 合肥工业大学学报(自然科学版), 2021, 44(8): 1139-1145. WANG R, ZHU C X, YUE J Y, et al. Isolation, identification and growth characterization of Microcystis aeruginosa in Chaohu Lake [J]. Journal of Hefei University of Technology (Natural Science), 2021, 44(8): 1139-1145(in Chinese).
[13] 张国维. 湖光岩玛珥湖溶解态氮与浮游植物及其氮吸收的研究[D]. 湛江: 广东海洋大学, 2014. ZHANG G W. Studies on dissovled nitrogen, phytoplankton and nitrogen uptake by phytoplankton in Huguangyan maar lake[D]. Zhanjiang: Guangdong Ocean University, 2014 (in Chinese).
[14] ABARIKE G A, SONG Z G, HAN Y Q, et al. Dissolved organic carbon concentration and its seasonal variation in the Huguangyan Maar Lake of Southern China[J]. Acta Geochimica, 2021, 40(5): 806-818. [15] 曹波. 铜绿微囊藻与小球藻种间竞争及其影响因素研究[D]. 广州: 暨南大学, 2010. CAO B. The interspecific competitions between Microcystis aeruginosa and Chlorella pyreniodosa and the factors affecting on this competition[D]. Guangzhou: Jinan University, 2010 (in Chinese).
[16] 马欠, 邓春暖, 郭锋锋. 温度对小球藻和铜绿微囊藻生长及叶绿素荧光特性的影响 [J]. 中州大学学报, 2018, 35(4): 108-112. MA Q, DENG C N, GUO F F. Effects of different temperature on growth of Chlorella and Microcystis meruginosa and chlorophyll fluorescence [J]. Journal of Zhongzhou University, 2018, 35(4): 108-112(in Chinese).
[17] ROBARTS R D, ZOHARY T. Temperature effects on photosynthetic capacity, respiration, and growth rates of bloom-forming cyanobacteria [J]. New Zealand Journal of Marine and Freshwater Research, 1987, 21(3): 391-399. doi: 10.1080/00288330.1987.9516235 [18] 金相灿, 李兆春, 郑朔方, 等. 铜绿微囊藻生长特性研究[J]. 环境科学研究, 2004, 17(S1): 52-54, 61. JIN X C, LI Z C, ZHENG S F, et al. Studies on the growth characteristics of Microcystis aeruginosa[J]. Research of Environmental Sciences, 2004, 17(Sup 1): 52-54, 61 (in Chinese).
[19] 陈建中, 刘志礼, 李晓明, 等. 温度、pH和氮、磷含量对铜绿微囊藻(Microcystis aeruginosa)生长的影响 [J]. 海洋与湖沼, 2010, 41(5): 714-718. CHEN J Z, LIU Z L, LI X M, et al. Effects of temperature, pH, nitrogen and phosphorus on growth of Microcystis aeruginosa [J]. Oceanologia et Limnologia Sinica, 2010, 41(5): 714-718(in Chinese).
[20] 张国维, 黄翔鹄, 李长玲, 等. 湖光岩玛珥湖叶绿素a与水质因子的相关分析及富营养化评价 [J]. 广东海洋大学学报, 2015, 35(1): 69-75. ZHANG G W, HUANG X H, LI C L, et al. Correlation analysis between Chlorophylla and water quality indices in Huguangyan maar lake and its eutrophication evaluation [J]. Journal of Guangdong Ocean University, 2015, 35(1): 69-75(in Chinese).
[21] 吴珊, 张晓萍, 张福萍. 2种藻类储磷释磷过程与生长情况对比 [J]. 河海大学学报(自然科学版), 2010, 38(1): 15-19. WU S, ZHANG X P, ZHANG F P. Comparative study on phosphorus reserve and release process and growth of two kinds of algae [J]. Journal of Hohai University (Natural Sciences), 2010, 38(1): 15-19(in Chinese).
[22] KIM H S, HWANG S J, SHIN J K, et al. Effects of limiting nutrients and N: P ratios on the phytoplankton growth in a shallow hypertrophic reservoir [J]. Hydrobiologia, 2007, 589(1): 317. doi: 10.1007/s10750-007-0727-1 [23] REDFIELD A C. The biological control of chemical factors in the environment [J]. Science Progress, 1960, 11: 150-170. [24] 贺艳艳, 李琴, 张瑜斌. 湖光岩玛珥湖浮游植物、浮游细菌生物量与可培养细菌数量的周年变化 [J]. 广东海洋大学学报, 2015, 35(1): 62-68. HE Y Y, LI Q, ZHANG Y B. Annual dynamics of phytoplankon biomass, bacterioplankton biomass and densities of culturable bacteria in Huguangyan maar lake [J]. Journal of Guangdong Ocean University, 2015, 35(1): 62-68(in Chinese).
[25] 张海涵, 王娜, 宗容容, 等. 水动力条件对藻类生理生态学影响的研究进展 [J]. 环境科学研究, 2022, 35(1): 181-190. ZHANG H H, WANG N, ZONG R R, et al. Research progress on influence of hydrodynamic conditions on algal physiology and ecology [J]. Research of Environmental Sciences, 2022, 35(1): 181-190(in Chinese).
[26] 李祥华. 水位下降引起的微弱扰动对浮游藻类生长的影响研究[D]. 重庆: 重庆大学, 2010. LI X H. Research on the influence of algae growth under feeble disturbance caused by water level drop[D]. Chongqing: Chongqing University, 2010 (in Chinese).
-
 点击查看大图
点击查看大图
计量
- 文章访问数: 3199
- HTML全文浏览数: 3199
- PDF下载数: 82
- 施引文献: 0
