

-
外来入侵植物通过自然或人为方式传播并定植在新的生态系统中,其生长或繁殖极大地影响生态系统的安全和本土物种多样性的保持[1-2];外来入侵植物在新的生态系统中竞争能力极强,能造成本土生态系统的崩溃,其防控和治理势在必行[3-4]。我国外来植物入侵形式严峻,现有外来入侵植物72科515种[5],其中1类恶性入侵物种34种[6],并且主要分布于我国东部沿海及西南地区。现针对入侵植物的研究主要集中在入侵植物特征、种类构成、入侵机理、防控方法等[7-8];并且有研究详细分析了不同区域或不同生态系统入侵植物的分布特征[9-10],这对于我国入侵植物管理和防控具有积极意义。我国西南内陆的四川盆地是外来植物入侵严重的地区,区域内分布着人口集中、资源交流频繁的城市生态系统,极易造成入侵植物扩散。但现阶段关于四川盆地城市生态系统外来入侵植物的相关研究匮乏,这对于四川盆地城市生态系统入侵植物的防控极其不利,因此亟待开展相关研究。
四川盆地位于我国西南,享有“天府之国”美誉[11],该区气候湿润,生物多样性丰富,水源涵养能力极强,对长江流域生态安全具有至关重要的作用[12]。四川盆地地理和环境特征明显:交通多样、道路密集,城市错落分布,资源交流频繁,外来入侵植物流通渠道多样;水、土资源丰富,外来入侵植物定植环境良好;四周丘陵高山环绕,中部平原肥沃,外来入侵植物可选择环境多样;因此该区极易成为外来入侵植物的重灾区。但四川盆地外来入侵植物现有的研究集中于园林类型[13],如紫茎泽兰入侵风险分析[14]等,城市外来入侵植物分布特征缺乏。因此以四川盆地城市典型生态系统为对象,开展野外植物群落调查,探究外来入侵植物在四川盆地城市典型生态系统中的分布特征,为四川盆地外来入侵植物的防控提供基础。
-
研究区域位于我国四川省,综合考虑其盆地城市的地理位置、人口分布、经济发展3大因子,选取成都、绵阳和眉山市作为四川盆地代表性城市类型。研究区域为典型的亚热带湿润季风气候,年降雨量1 200 mm左右,年均气温16 ℃左右,气候温暖湿润。研究区域海拔250 ~ 700 m,主要由平原和低山丘陵组成,地势开阔,密布河流、湖泊,土壤主要由紫色土、黄壤和黄棕壤组成;区域森林覆盖率高,农业发达,是我国水稻和油菜籽的主要产区。
-
根据四川盆地城市生态系统特征,初步将城市生态系统划分为陆地旱地和湿地两大类型,最后综合典型城市的环境、生态等特征,将旱地生态系统划分为人为干扰较重的道路生态系统(包含公园、草坪和人工林等);人工遗弃后干扰较少的荒地生态系统(包含建筑荒地、绿化荒地等);因四川盆地城市地理区划中长期分布着人为干扰较重的农用田地(农用田地、自留菜地),也是四川盆地城市生态系统中的重要组成部分。将湿地生态系统划分为大小不一的湖泊生态系统,常年流水且具有周期性滩涂的河流生态系统。道路、荒地、农田、湖泊、河流5个生态系统用于开展植物样方调查和植物资源统计,了解外来入侵植物分布特征与现状。
-
在研究区域,道路、荒地和农田生态系统按照陆地植物调查方法进行样方调查,湖泊主要调查水深30 cm内和邻水(3 m内)区域的植物,河流生态系统主要调查滩涂与河岸(2 m内)区域的植物[15]。于2019年6月和2020年6月,确定区域内代表生态系统的地理位置,在每个调查样本区域,进行1次样本调查,在调查区域内选择3块5 m × 5 m的大样方,在其中选择典型1 m × 1 m 的小样方进行调查。调查小样方中的草本、藤本、灌木、小乔木和乔木幼苗,排除高大乔木。记录样方中各物种的数量、高度、多度、盖度,如果样方中的藤本或匍匐草本盖度占据本样方面积的10%以上,则所有植物只统计相对盖度(占整个样方的比值),作为样方内植物物种多样性计算的基础值。
-
依据《中国高等植物彩色图鉴》[16]和《四川植物志》[17]确定所有物种的详细信息,并且根据中国植物http://www.iplant.cn/与《中国入侵植物名录》来对比和确定所调查的物种的分布、转移历史,确定物种是否为外来入侵物种,以及入侵物种等级。
-
在确定后的样方内调查植物物种和分布情况,对分布的丰富度指数(Species richness, R)、生态优势度指数(Simpson diversity index, D)、物种多样性指数(Shannon-Wiener index, H′)和均匀度指数(Pielou index, E)进行分析,明确各群落植物生物多样性。群落物种丰富度指数[18],见式(1)。
式(1)中,R为群落物种丰富度指数,S为群落中的总物种数。生态优势度指数[19],见式(2)。
式(2)中,D为Simpson指数,N为群落中全部种的总个体数(总盖度),Ni为第i种的个体数(单个物种在群落中的盖度),S为群落中的总物种数。物种多样性指数(Shannon-Wiener指数)[20],见式(3)。
式(3)中,H′为Shannon-Wiener指数,Pi =Ni /N,Pi为群落中第i种个体数占群落中全部物种总个体数的比例(单个物种在群落中的盖度占总盖度的比例),S为群落中的总物种数,均匀度指数(Pielou指数)[19],见式(4)。
式(4)中,E为Pielou指数。所有样方中,如果无外来入侵植物,则样方记为无入侵种;具有外来入侵植物,则记为具入侵种。各系统中的外来植物入侵率=(具入侵样本量/调查样本总量)×100%;各入侵植物占群落比=各入侵物种个体数(盖度)/群落内总植物个体数(总盖度)。对各典型入侵植物在不同生态系统中占比差异,如入侵植物的样本量少于3个,则不进入统计。
-
所有数据为均值加标准误(ME±SE),利用单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)分析各生态系统中植物物种多样性之间的差异,具入侵植物和无入侵植物在各生态系统中植物物种多样性之间的差异,不同外来入侵植物在相同生态系统中的占比差异,各典型入侵植物在不同生态系统中的占比差异,显著水平为P < 0.05。所有数据利用Spss 19.0进行分析,Origin 9.0作图。
-
四川盆地城市典型生态系统外来植物入侵比例极高,各生态系统中外来植物入侵比例都超过60%,见表1。
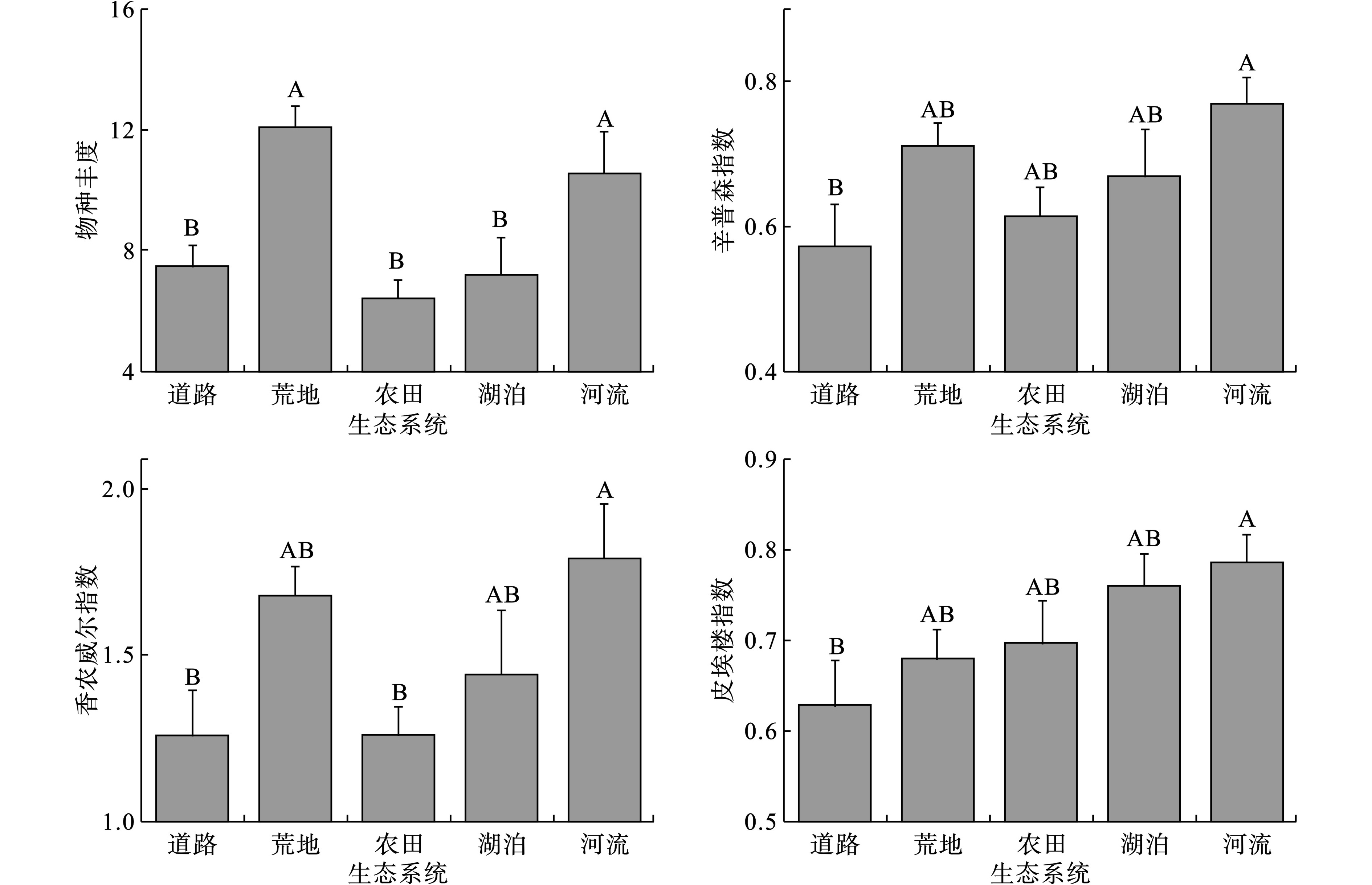
在5种典型的生态系统中,人为干扰较少的荒地与河流生态系统中外来植物入侵率达到100%,人为干扰较大的道路和农田生态系统中入侵率分别达到64.29%和72.73%。调查到的外来入侵物植物共计20种,分布于11科,18属,见表2。
喜旱莲子草(Alternanthera philoxeroides)是所有生态系统中共有的外来入侵植物;除湖泊生态系统外,小蓬草(Erigeron canadensis)在其他4个生态系统中都有分布;除农田生态系统外,白车轴草(Trifolium repens)在其他4个生态系统中都有分布。道路生态系统中,外来入侵植物白车轴和喜旱莲子草分布最广。荒地生态系统中,外来入侵植物种类最多,达12种,入侵植物小蓬草分布最广。农田生态系统中,外来入侵植物小蓬草分布最广。湖泊生态系统中,外来入侵植物喜旱莲子草分布最广。河流生态系统中,外来入侵植物喜旱莲子草和白车轴草分布最广。
-
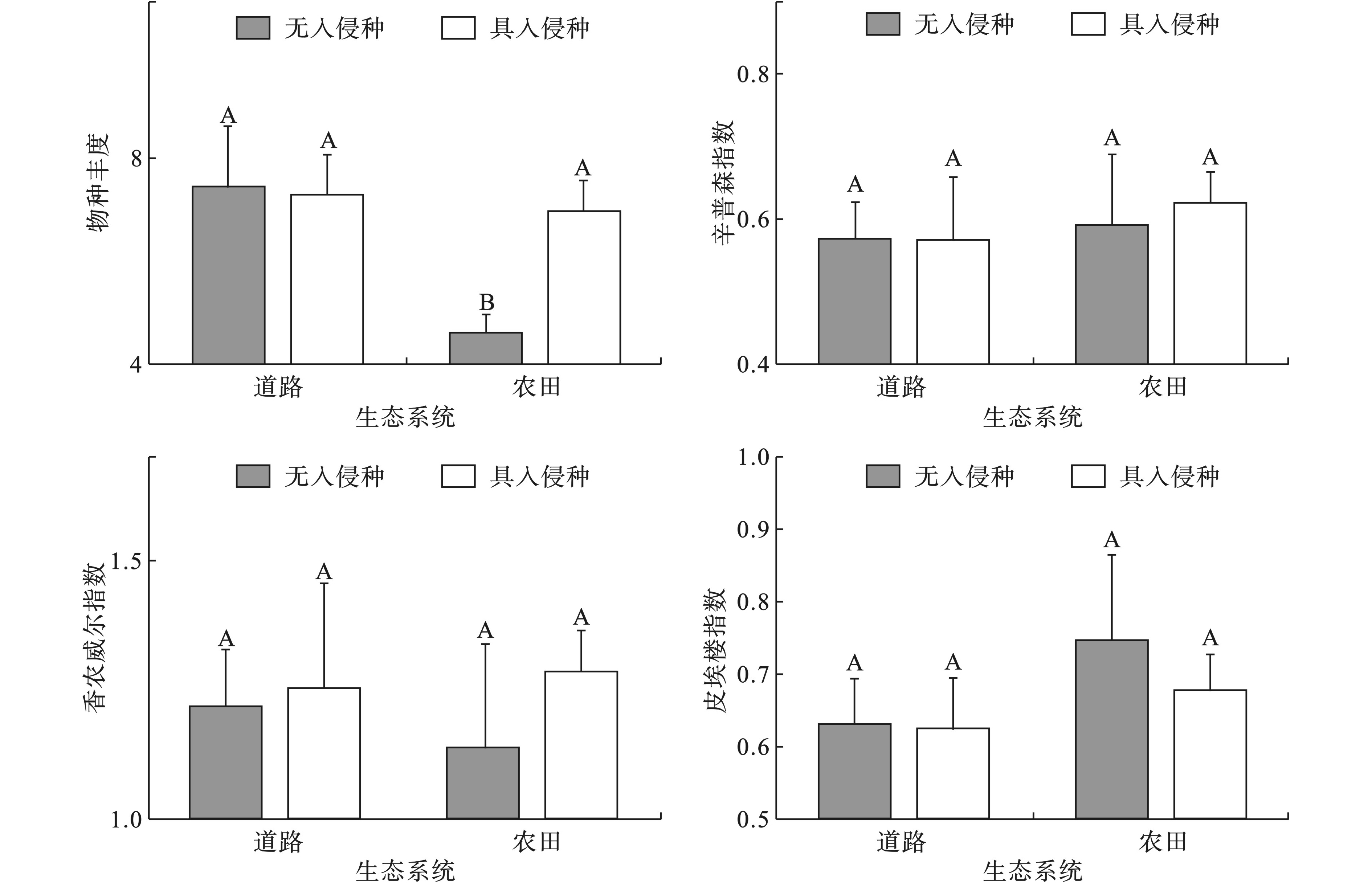
四川盆地城市5种典型生态系统类型中物种多样性分布差异较大,植物分布均匀度不一,见图1。道路和农田生态系统中具入侵和无入侵植物群落物种多样性,见图2。
5种生态系统中,荒地、湖泊生态系统全部具有外来植物入侵,河流生态系统无入侵植物的样本量极少,因此,3种生态系统类型不进行多样性比较。在农田生态系统中,无入侵和具入侵植物的群落物种多样性存在显著性差异(P < 0.05),具入侵植物的群落物种多样性和无入侵植物的群落物种丰度差异显著,无入侵植物的群落物种丰度显著小于具入侵植物的群落;而辛普森指数、香农威尔指数、均匀度指数却相似,并没有显著差异。
-
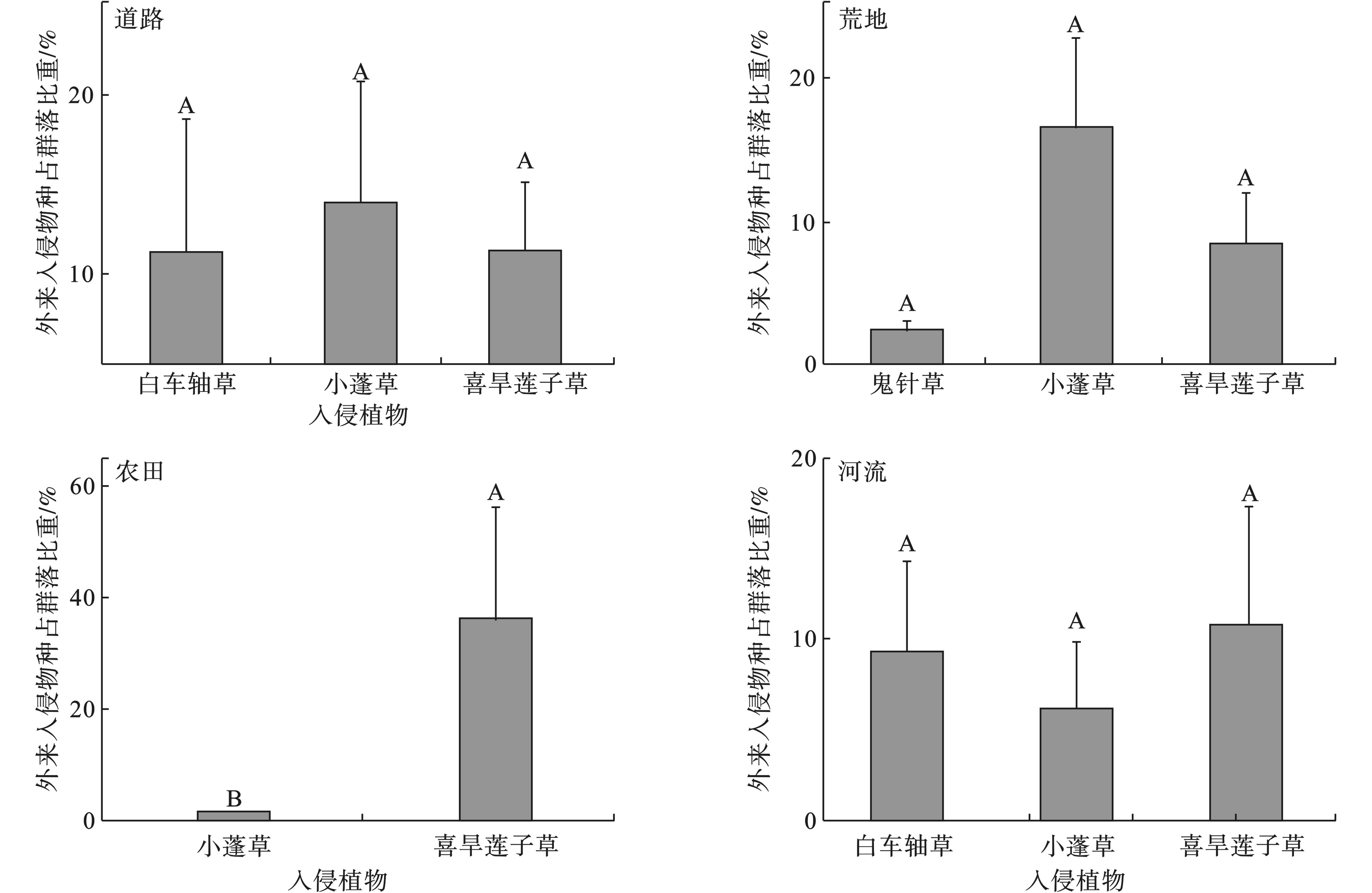
不同生态系统中,各外来入侵植物在群落中占比存在差异,见图3。道路生态系统中,典型的入侵物种为白车轴草、小蓬草和喜旱莲子草,3个入侵植物在道路生态系统中植物群落中占比都超过10%,且小于15%。荒地生态系统中,鬼针草、小蓬草和喜旱莲子草为典型入侵植物,小蓬草占植物群落比例最高,达15%,而最低的鬼针草占比小于5%。农田生态系统中,典型的入侵物种为小蓬草和喜旱莲子草,小蓬草占群落比例小于5%,但喜旱莲子草占群落比例超过30%,两种草本在农田生态系统中占群落比例差异显著。河流生态系统中,典型的入侵植物为白车轴草、小蓬草和喜旱莲子草,3种入侵草本在群落中的占比分布于6%至11%,水平相当。湖泊生态系统中典型的入侵植物只有喜旱莲子草,未进行种间占比差异分析。
荒地生态系统中,群落物种丰度高达12.1,而最低的农田生态系统物种丰度只有6.45;荒地、河流生态系统的物种丰度比道路、农田、湖泊生态系统的物种丰度高,且具有显著性差异(P < 0.05)。5种生态系统中,河流生态系统中植物物种多样性(辛普森指数和香农威尔指数)最高,而道路生态系统中植物物种多样性最低,且两个生态系统中植物物种多样性具有显著性差异(P < 0.05)。在所有生态系统中,河流生态系统中植物群落中物种分布最均匀,而道路生态系统中植物分布均匀度最低,且两种生态系统中植物群落均匀度具有显著性差异(P < 0.05)。5种生态系统的物种多样性并没有呈现湿生和旱生的两级分化,但是均匀度指数表明河流生态系统中物种分布较为均匀。
5种生态系统中广布的典型入侵植物为小蓬草和喜旱莲子草,两种入侵植物在不同生态系统中占植物群落比例分布不一。喜旱莲子草在5种生态系统中占群落比例出现了明显的差异,农田的36.75%和荒地的8.5%之间差异显著(P < 0.05);农田中的占比极大值和极小值之间的差值较大,表3。
-
四川省外来入侵植物高达155种,本次调查的四川盆地城市典型生态系统中外来入侵植物多达20种,分布于菊科(Asteraceae)、豆科(Leguminosae)、苋科(Amaranthaceae)和十字花科(Brassicaceae)等。四川盆地生态环境适宜于外来植物定植[21];并且四川盆地城市典型生态系统物质交流过程造成大量外来植物被作为景观资源引入本土生态系统中,如白车轴草、红花酢浆草等,因此造成了四川盆地外来植物种类丰富[22]。大量外来植物可能对乡土植物的生存空间造成极大的冲击,如竞争能力较弱的黄鹌菜(Youngia Japonica)、野大豆(Glycine soja)和海金沙(Lygodium japonicum)等本土种,极易从生态系统中消退。5种典型生态系统中外来入侵植物具有明显的分布特征,但还需要更加细致和深入的调查,以得到更加详细的外来入侵植物类型和特征资料。
外来植物对四川盆地城市5种生态系统入侵率超过60%;其中人为干扰较少的荒地与河流生态系统中,外来植物入侵率已经高达100%,这些结果与金佛山地区外来入侵植物分布情况差异较大[23]。这可能是由于四川盆地城市生态系统的特性,资源交流带来的植物入侵可能已经蔓延到整个城市生态系统,而人工除草、防治等干预行为反而减少了入侵植物,人为干扰防除了部分入侵植物。在5种生态系统中,人为干扰较大的道路、农田生态系统的物种多样性低于人为干扰较小的荒地、河流生态系统,人为耕作的农田系统中入侵物种增加了生态系统物种多样性,人为干扰可能影响了乡土植物和外来入侵植物的分布[24],这些都说明人为干扰可能是影响外来植物定植和扩散的重要因素。在其他研究中,人为干扰被证明可以有效防治外来植物入侵[25]。因此,四川盆地城市外来入侵植物可以积极采用人为拔除、养护等方式来进行防治。
四川盆地城市中分布最广的外来入侵物种是白车轴草、小蓬草和喜旱莲子草。喜旱莲子草和小蓬草都是来自于南美洲热带/亚热带地区的1级恶性入侵草本,两种草本的强入侵能力迅速占据生态位,成为生态系统内的优势物种,特别是小蓬草繁殖能力极强,扩散迅速[26]。白车轴草原产于欧洲和北非,属于2级严重入侵草本。小蓬草和喜旱莲子草在生态系统内的盖度最高分别达到了41.84%和70.42%,可能极大的影响了本土物种的正常生长和繁殖过程[27];并且两种草本分布广泛,已经逸生为四川盆地城市中的野生物种,可能对乡土植物群落产生较大的影响。
综合四川盆地城市典型生态系统外来入侵植物的分布特征,初步提出防治对策:继续深入完善四川盆地城市生态系统中外来入侵植物数据库,明确外来入侵植物的类型、分布状态、生物学和生态学特性,建立动态数据库和危害评估体系;加大人工防治力度,通过人工拔除、人工管护、源头管控和截断扩散渠道等方式逐步治理外来入侵植物,使外来入侵植物防治进入常态化;建立并强化外来植物准入机制,严格把控外来植物引种过程,坚决避免有害植物引入。
-
四川盆地典型城市生态系统中,5个典型生态系统中外来植物入侵严重,人为干扰较小的荒地与河流生态系统中入侵率高达100%。样方调查发现四川盆地城市外来入侵植物有11科18属20种;喜旱莲子草、小蓬草、白车轴草为广泛分布的外来入侵植物;喜旱莲子草和小蓬草是主要的恶性入侵物种,并且已经逸生为野生物种。外来植物入侵显著影响了植物群落物种多样性和占比。人为干扰能够影响外来入侵植物的分布,需要建立更加严格的防治对策,以应对四川盆地城市生态系统外来入侵植物问题。物问题。
城市典型生态系统外来入侵植物分布特征
——以四川盆地为例Distribution characteristic of invasive plants in typical urban ecosystem ——A case study of Sichuan Basin
-
摘要: 在四川盆地城市生态系统中,选择最典型的道路、荒地、农田、湖泊、河流5个生态系统对外来植物进行调查,分析外来入侵植物组成和分布特征。结果表明:四川盆地城市典型生态系统外来植物入侵形势严峻,河流和荒地生态系统外来植物入侵率高达100%;5个典型生态系统中有外来入侵植物共20种分布于11科18属;喜旱莲子草和小蓬草是分布最广的恶性入侵物种,并且已经逸为野生。外来入侵植物对四川盆地城市生态系统物种分布产生极大影响,但人为干扰能够显著影响外来入侵物种的分布。最后提出四川盆地城市生态系统外来入侵植物防治措施。Abstract: The quadrat survey method was used to analyze the vegetation community and distribution characteristic of invasive plants in the urban ecosystem of Sichuan Basin. The road, wasteland, farmland, lake and river ecosystem were selected as the typical ecological systems. Results showed that the alien plants invaded the city ecosystem of Sichuan Basin seriously with an invaded ratio 100% encroached river and wasteland ecosystems. There were 20 species alien plants which distributed in 11 families and 18 genera for the five typical ecosystems. Alternanthera philoxeroides and Erigeron Canadensis were the most widely serious invasive species, and the two invasive plants had been escaped and became the wild plant. The invade exotic plant impact seriously on the plant distribution in urban ecosystem of Sichuan Basin. However, the artificial disturbance could affect distribution of the invasive plants. The control strategies of the invasive plants in the urban ecosystem were also proposed.
-
Key words:
- southwest China /
- biosafety /
- ecological type /
- biodiversity
-

-
表 1 5种生态系统中外来植物入侵率
生态系统 调查样本总量 具入侵植物样本量 入侵率/% 道路 14 9 64.29 荒地 10 10 100.00 农田 11 8 72.73 湖泊 6 5 83.33 河流 7 7 100.00  下载: 导出CSV
下载: 导出CSV
表 2 入侵植物的入侵等级、原产地、分布量
种 科 属 入侵等级 原产地 道路 荒地 农田 湖泊 河流 分布量 分布量 分布量 分布量 分布量 白车轴草
Trifolium repens豆科
Leguminosae车轴草属
Trifolium2 北非、中亚、
西亚和欧洲5 1 / 1 4 黑麦草
Lolium perenne禾本科
Gramineae黑麦草属
Lolium4 欧洲 1 / / / / 小蓬草
Erigeron canadensis菊科
Compositae飞蓬属
Erigeron1 北美洲 4 8 5 / 3 喜旱莲子草
Alternanthera philoxeroides苋科
Amaranthaceae莲子草属
Alternanthera1 巴西 5 4 3 4 4 问荆
Equisetum arvense木贼科
Equisetaceae木贼属
Equisetum6 (建议排除) 1 / / / / 细叶旱芹
Cyclospermum leptophyllum伞形科
Umbelliferae细叶旱芹属
Cyclospermum4 南美洲 1 1 / / / 刺槐
Robinia pseudoacacia豆科
Leguminosae刺槐属
Robinia3 北美洲 / 1 / / / 鬼针草
Bidens pilosa菊科
Compositae鬼针草属
Bidens1 美洲 / 3 2 / / 红花酢浆草
Oxalis corymbosa酢浆草科
Oxalidaceae酢浆草属
Oxalis4 热带美洲 / 1 / 1 / 灰绿藜
Chenopodium glaucum藜科
Chenopodiaceae藜属
Chenopodium4 不详 / 1 / / / 芥菜
Brassica juncea十字花科
Cruciferae芸苔属
Brassica6 (建议排除) / 2 / / / 牛膝菊
Galinsoga parviflora菊科
Compositae牛膝菊属
Galinsoga2 南美洲 / 1 / / / 荠
Capsella bursa-pastoris十字花科
Cruciferae荠属
Capsella4 西亚和欧洲 / 1 1 1 / 紫苜蓿
Medicago sativa豆科
Leguminosae苜蓿属
Medicago4 西亚 / 1 / / / 欧洲千里光
Senecio vulgaris菊科
Compositae千里光属
Senecio4 欧洲 / / 1 / / 苏门白酒草
Erigeron sumatrensis菊科
Compositae飞蓬属
Erigeron1 南美洲 / / 1 / / 五爪金龙
Ipomoea cairica旋花科
Convolvulaceae番薯属
Ipomoea1 可能为美洲 / / 1 / / 钻叶紫菀
Aster subulatus菊科
Compositae紫菀属
Aster1 北美洲 / / 1 / / 蓖麻
Ricinus communis大戟科
Euphorbiaceae蓖麻属
Ricinus2 东非 / / / / 1 野胡萝卜
Daucus carota伞形科
Umbelliferae胡萝卜属
Daucus2 欧洲 / / / / 2 注:“/” 表示该物种在该生态类型下无分布。
下载: 导出CSV
表 3 不同生态系统中小蓬草和喜旱莲子草在植物群落内的占比
% 物种 生态系统 占植物群落比 极小值 极大值 小蓬草 Erigeron canadensis 荒地 16.48±6.28A 0.66 41.84 道路 13.9±6.8A 1.94 32.48 农田 1.66±0.63A 0.35 3.57 河流 6.28±3.61A 1.61 13.39 喜旱莲子草 Alternanthera philoxeroides 道路 11.34±3.76AB 0.93 20.00 荒地 8.5±3.45B 2.75 17.31 农田 36.75±19.69A 2.22 70.42 湖泊 26.95±6.98AB 12.22 40.00 河流 10.91±6.41AB 2.56 30.00 注:不同大写字母表示各入侵植物在不同生态系统之间占植物群落比例差异显著。
下载: 导出CSV
-
[1] MOONEY H A, CLELAND E E. The evolutionary impact of invasive species[J]. Proceedings of the National Academy of Sciences of the United States of America, 2001, 98: 5446 − 5451. doi: 10.1073/pnas.091093398 [2] ZHENG Y L, FENG Y L, VALIENTE-BANUET A, et al. Are invasive plants more competitive than native conspecifics? Patterns vary with competitors[J]. Scientific Reports, 2015, 5: 15622. doi: 10.1038/srep15622 [3] EHRENFELD J G. Effect of exotic plant invasion on soil nutrient cycling processes[J]. Ecosystems, 2003, 6: 503 − 523. doi: 10.1007/s10021-002-0151-3 [4] SHIFERAW H, SCHAFFNER U, WOLDEAMLAK B, et al. Modelling the current fractional cover of an invasive alien plant and drivers of its invasion in a dryland ecosystem[J]. Scientific Reports, 2019, 9(1): 1 − 12. [5] 闫小玲, 刘全儒, 寿海洋, 等. 中国外来入侵植物的等级划分与地理分布格局分析[J]. 生物多样性, 2014, 22(5): 667 − 676. [6] 马金双. 中国入侵植物名录[M]. 北京: 高等教育出版社, 2014. [7] 吴晓雯, 罗晶, 陈家宽, 等. 中国外来入侵植物的分布格局及其与环境因子和人类活动的关系[J]. 植物生态学报, 2006, 30(4): 576 − 584. doi: 10.3321/j.issn:1005-264X.2006.04.006 [8] 马瑞燕, 王韧, 丁建清. 利用传统生物防治控制外来杂草的入侵[J]. 生态学报, 2003, 23(12): 186 − 197. [9] 王苏铭, 张楠, 于琳倩, 等. 北京地区外来入侵植物分布特征及其影响因素[J]. 生态学报, 2012, 32(15): 4618 − 4629. [10] 任颖, 何萍, 徐杰, 等. 滦河流域河岸带入侵植物分布特征及其与环境的关系[J]. 应用生态学报, 2017, 28(6): 1843 − 1850. [11] 袁廷栋. "天府之国"由来的历史考察[J]. 社会科学研究, 1985(1): 54 − 59. [12] 洪步庭, 任平, 苑全治, 等. 长江上游生态功能区划研究[J]. 生态与农村环境学报, 2019, 35(8): 1009 − 1019. [13] 何兵, 崔莉, 宋丽娟, 等. 成都园林入侵植物的调查及区系分析[J]. 西南农业学报, 2011, 24(5): 312 − 317. [14] 田茂洁. 川西南紫茎泽兰(eupatorium adenophorum)旱季光合生理及其在四川盆地的入侵风险分析[J]. 西华师范大学学报:自然科学版, 2011, 31(4): 42 − 49. [15] 方精云, 王襄平, 沈泽昊, 等. 植物群落清查的主要内容、方法和技术规范[J]. 生物多样性, 2009, 17(6): 533 − 548. [16] 中国高等植物彩色图鉴编委. 中国高等植物彩色图鉴[M]. 北京: 科学出版社, 2016. [17] 四川植物志编辑委员会. 四川植物志[M]. 成都: 四川人民出版社, 1981. [18] 尚玉昌. 普通生态学 (第三版)[M]. 北京: 北京大学出版社, 2010. [19] PIELOU E C. Mathematical ecology[M]. New York: John Wiley and Sons Incorporation, 1975. [20] MAGURRAN A E. Ecological diversity and its measurement[M]. New Jersey: Princeton University Press, 1988. [21] 冯建孟, 董晓东, 徐成东. 中国外来入侵植物区系组成的大尺度格局及其气候解释[J]. 植物科学学报, 2009, 27(2): 159 − 164. doi: 10.3969/j.issn.2095-0837.2009.02.006 [22] TILMAN D, LEHMAN C. Human-caused environmental change: Impacts on plant diversity and evolution[J]. Proceedings of the National Academy of Sciences of the United States of America, 2001, 98(10): 5433 − 5440. doi: 10.1073/pnas.091093198 [23] 孙娟, 杨国锋, 陈玉成, 等. 金佛山自然保护区外来入侵植物种及其分布情况[J]. 草业学报, 2009, 18(3): 36 − 44. [24] 朱枫, 王四海, 陈剑, 等. 外来入侵植物肿柄菊在云南的分布特征及其影响因素[J]. 生态学杂志, 2018, 37(9): 2573 − 2580. [25] 邵志芳, 赵厚本, 邱少松, 等. 深圳市主要外来入侵植物调查及治理状况[J]. 生态环境学报, 2006, 15(3): 587 − 593. doi: 10.3969/j.issn.1674-5906.2006.03.029 [26] ZAPLATA M K, WINTER S, BIEMELT D, et al. Immediate shift towards source dynamics: The pioneer species Conyza canadensis in an initial ecosystem[J]. Flora, 2011, 206(11): 928 − 934. doi: 10.1016/j.flora.2011.07.001 [27] FAN S F, YU D, LIU C H. The Invasive Plant Alternanthera philoxeroides was suppressed more intensively than its native congener by a native generalist: Implications for the biotic resistance hypothesis[J]. PLoS ONE, 2013, 8(12): e83619. doi: 10.1371/journal.pone.0083619 -
 点击查看大图
点击查看大图
图( 3) 表( 3)
计量
- 文章访问数: 4434
- HTML全文浏览数: 4434
- PDF下载数: 42
- 施引文献: 0
