


-
浮游植物是湖泊生态系统重要的初级生产者,在湖泊生态系统的能量流动和物质循环中发挥着重要的作用[1]。在自然水体中,浮游植物的种类、数量和分布易受到光照、水质以及扰动等外界环境因素的影响,会随环境因素的变化而变化,因此,浮游植物被广泛应用于水域的水质评价[2-4]。
太湖是中国五大淡水湖之一,全湖水域面积达2 338 km2,是一个典型的浅水湖泊[5]。从20世纪80年代中后期开始,太湖流域周边经济快速发展,各种点源、面源污染物不断排入太湖中,湖水富营养化问题日益严重,每年的5~11月,蓝藻水华频频发生,已经严重危害居民用水安全[6-7]。
2007年太湖蓝藻水华事件以后,国家和地方政府投入了大量的资金进行太湖周边污染物的控制以及太湖水治理,太湖水质整体明显改善[8-9]。然而,赵凯[10]研究发现,2015年太湖水生植被的严重退化,其中2015年夏季水生植被分布区面积仅占2014年夏季的20.14%,2015年冬季水生植被分布区面积仅占2014年冬季的8.94%。太湖水生植物的变化反映了太湖水质和浮游植物群落结构的变化特征。为了更好的了解目前太湖的水质情况,2017年7月至2018年6月对太湖的梅梁湾(藻型湖区)和东太湖(草型湖区)浮游植物群落结构和水质进行采样调查。其中,梅梁湾是典型的藻型湖区,同时是太湖中富营养化最严重的湖区之一,每年都会爆发大规模的蓝藻水华;而东太湖则是草型湖区,湖中水生植物较多,富营养化和蓝藻水华问题较轻。通过周年采样调查,比较分析太湖2个典型湖区的浮游植物群落结构变化特征,进一步了解目前太湖的水质状况变化,为太湖的管理提供科学依据。
-
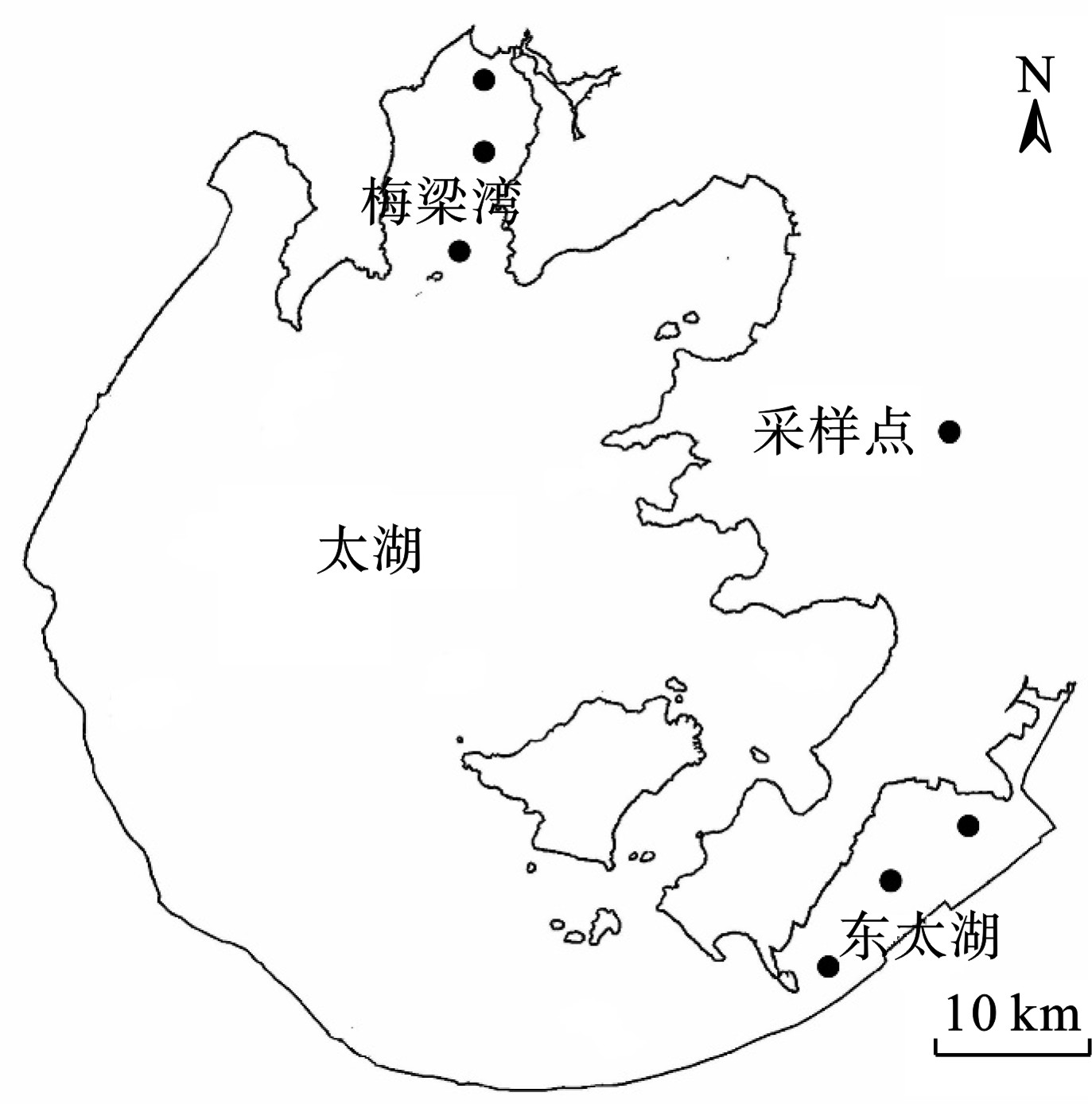
2017年7月至2018年6月,在梅梁湾和东太湖布设6个采样点,进行浮游植物和水体环境因子的现场调查,见图1。
-
样品每月采集1次,用5 L的有机玻璃柱状采水器分2层采集湖水,分别为表层(0~50 cm)和中下层(100~150 cm),然后混匀,取1 L水样加入鲁哥试剂(最终浓度1%)固定保存浮游植物。另取1 L混合水样,带回实验室后按文献[11]方法分析水样理化指标(总氮、溶解性总氮、氨氮、硝氮、总磷、溶解性总磷、正磷酸盐、COD和悬浮颗粒物)和叶绿素a(Chl-a)浓度,并在现场测定水深、水温、透明度和pH,水样理化指标测定方法见文献[11]。
-
将1 L已加入鲁哥试剂的水样带回实验室后静置48 h,去掉上清液将其浓缩为50 mL,即为最终保存样品。在显微镜(Olympus Corporation, Tokyo, Japan)400×下进行分类计数,每个样品每次计数100个视野,计数至少3次,取其平均值作为最后结果,并按公式计算实际浮游植物数量。浮游植物的鉴定参考《中国淡水藻类——系统、分类及生态》[12]。浮游植物数量换算公式,见式(1)。
式(1)中:N为每升水样中浮游植物数量,cells/L;A为计数框面积,mm2;Vs为1 L水样浓缩后的体积,mL;n为计数所得浮游植物个数;Ac为计数面积,mm2;Va为计数框体积,mL。
浮游植物的生物量由不同种类浮游植物的单个生物量乘以细胞个数计算所得[13],本研究中浮游植物生物量均为湿重。
浮游植物群落的生物多样性采用Shannon-Wiener生物多样性指数(H)表征,见式(2~3)。
式(2~3)中:
$ {n_i} $ 为浮游植物群落中第$ i $ 物种的个体数;$ N $ 为所有浮游植物物种的个体数之和;$ S $ 为浮游植物群落中物种总数;$ {P_i} $ 为第$ i $ 种的个体数占总个体数的比例。 -
本文中图使用Origin 2017软件绘制,用t检验法分析两个湖区物理、化学和浮游植物数据的差异,所有的数据均使用SPSS 22.0软件进行统计分析,采用P<0.05作为显著性差异水平。
-
本研究梅梁湾和东太湖环境因素相关值,见表1。
表1可知,梅梁湾富营养水平高于东太湖,其中,梅梁湾湖区的总氮、总磷、溶解性总氮、溶解性总磷都显著高于东太湖湖区(P < 0.05)。
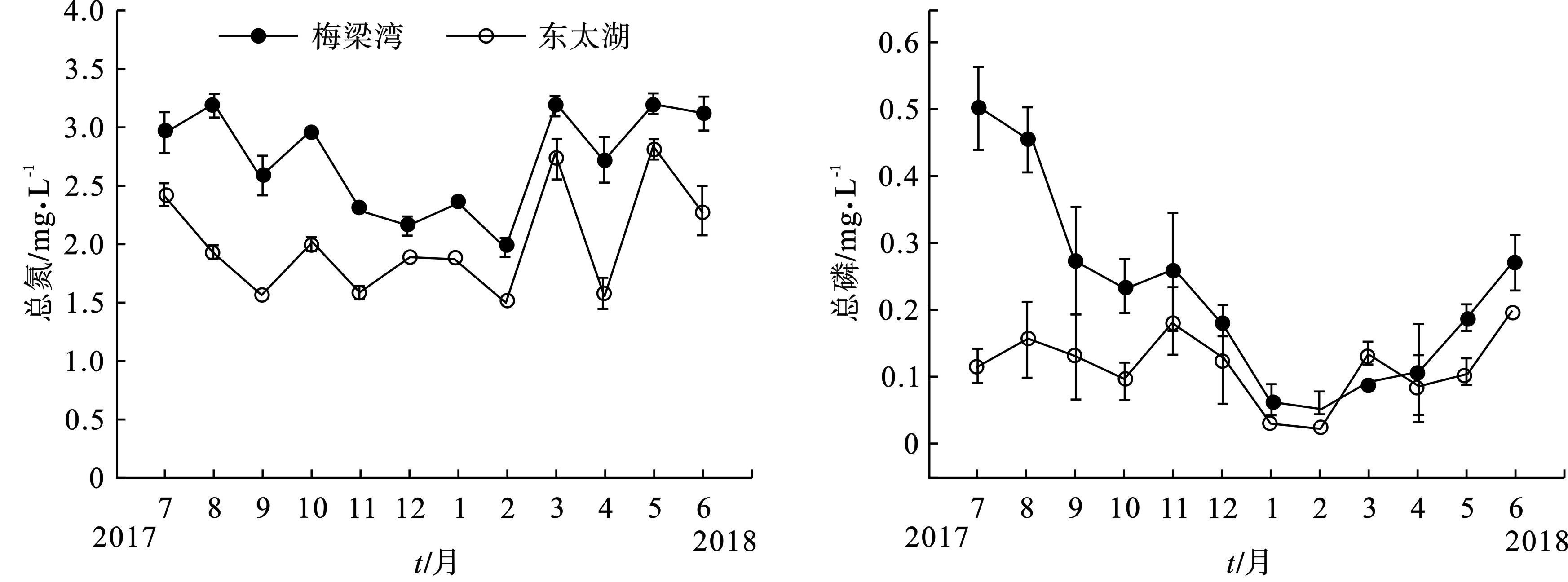
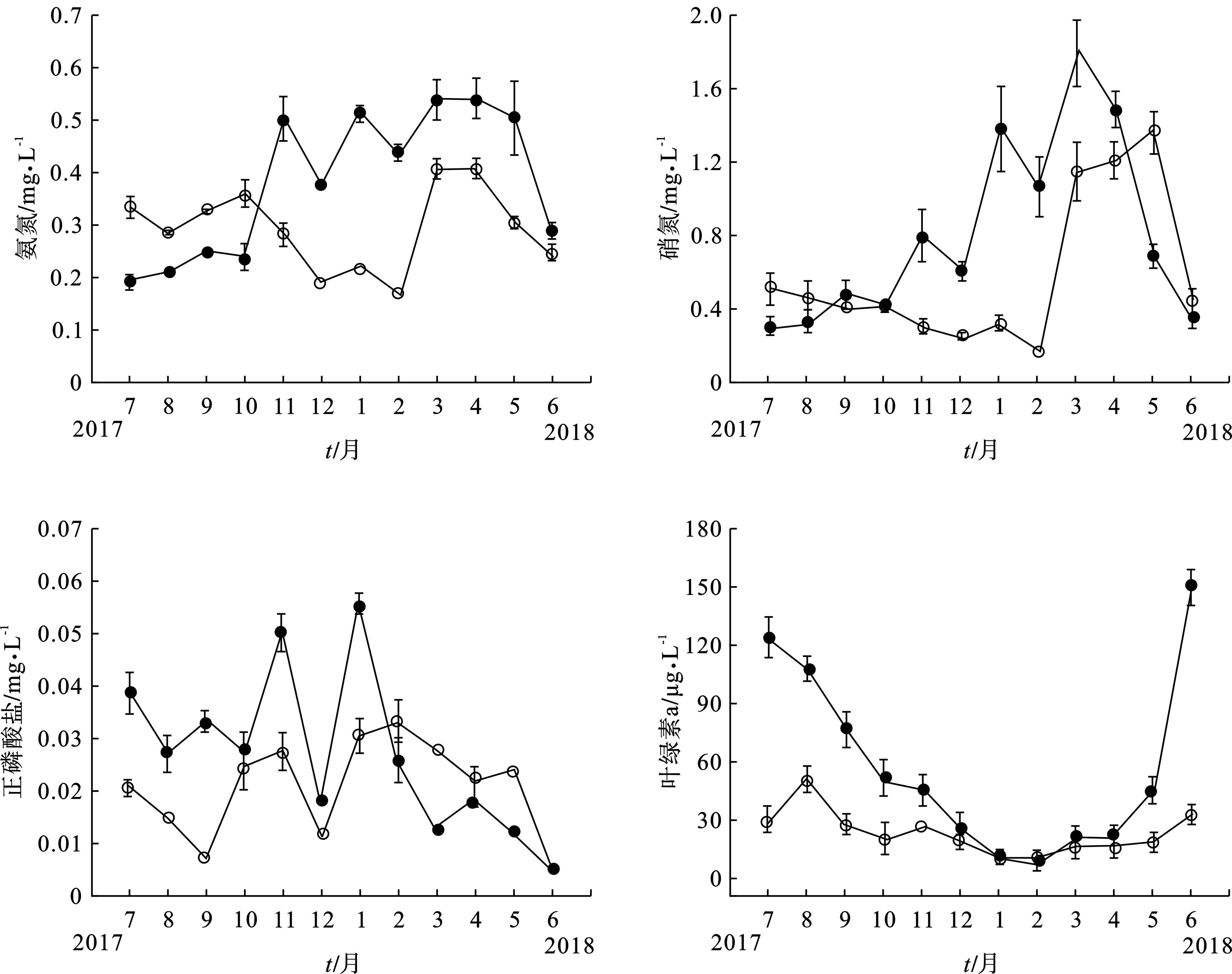
主要环境参数的季节变化趋势,见图2a~e。梅梁湾总氮最高值和最低值分别出现在2018年5月和2018年2月,分别为3.200、1.988 mg/L;东太湖总氮最高值和最低值也分别出现在2018年5月和2018年2月,其值分别为2.827和1.523 mg/L,见图2a。东太湖、梅梁湾总磷最大值分别为0.203、0.505mg/L,分别出现在2018年6月和2017年7月,最小值分别为0.023、0.052 mg/L,都出现在2018年2月,见图2b。
氨氮和硝氮浓度总体呈现出梅梁湾湖区高于东太湖湖区的特征,见图2c~d。东太湖、梅梁湾湖区的正磷酸盐浓度之间没有发现明显的规律性,见图2e。从图2f可见,梅梁湾叶绿素a 浓度显著高于东太湖,尤其在2018年6月,东太湖叶绿素a质量浓度为32.61 μg/L,而梅梁湾却高达150.57 μg/L。
-
东太湖、梅梁湾分别出现37和39种浮游植物。2个湖区有相同的浮游植物32种,只出现在梅梁湾的有尖尾蓝隐藻Chroomonas acuta、星杆藻Asterionella sp.、密集锥囊藻Dinobryon sertularia、囊裸藻Trachelomonas spp.、微小四角藻Tetraedron minimum、河生集星藻Actinastrum fluviatile、美丽网球藻Dictyosphaerium pulchellum;只出现在东太湖的物种有颤藻Oscillatoria spp.、四角十字藻Crucigenia quadrata、三角四角藻Tetraedron trigonum、肥壮蹄形藻Kirchneriella
obesa和柯氏并联藻Quadrigula chodatii。 -
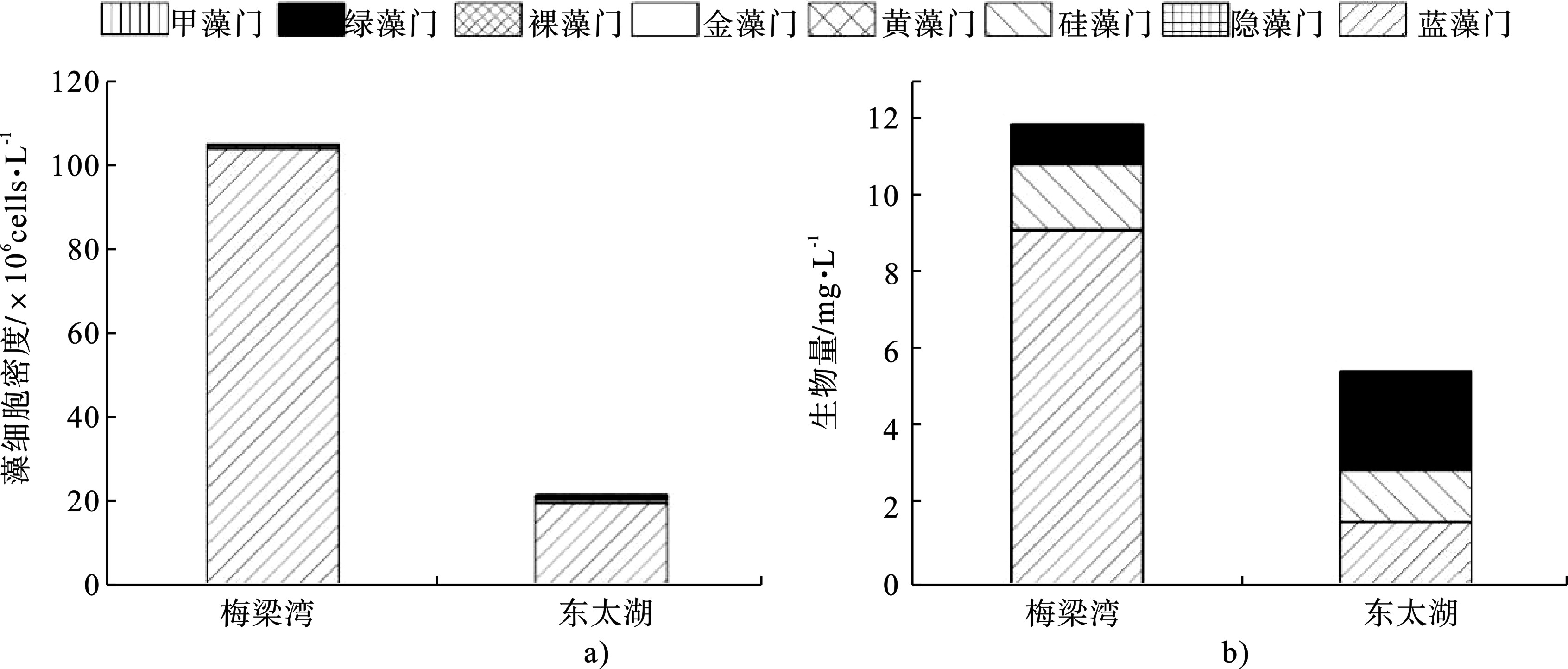
梅梁湾和东太湖的浮游植物年平均密度分别为104.73×106和21.18×106 cells/L,见图3a,前者显著高于后者(P<0.01),并且2个湖区蓝藻门均占主要部分,分别占总浮游植物的98.78%和90.89%。
梅梁湾浮游植物生物量为11.84 mg/L,东太湖为5.49 mg/L,前者显著大于后者(P<0.01),见图3b。梅梁湾湖区依然是蓝藻生物量占大多数,为76.96%,绿藻和硅藻占小部分,分别为8.48%和14.16%,见图3b。东太湖湖区绿藻生物量占46.34%,蓝藻和硅藻生物量分别为29.38%和24.13%,见图3b。
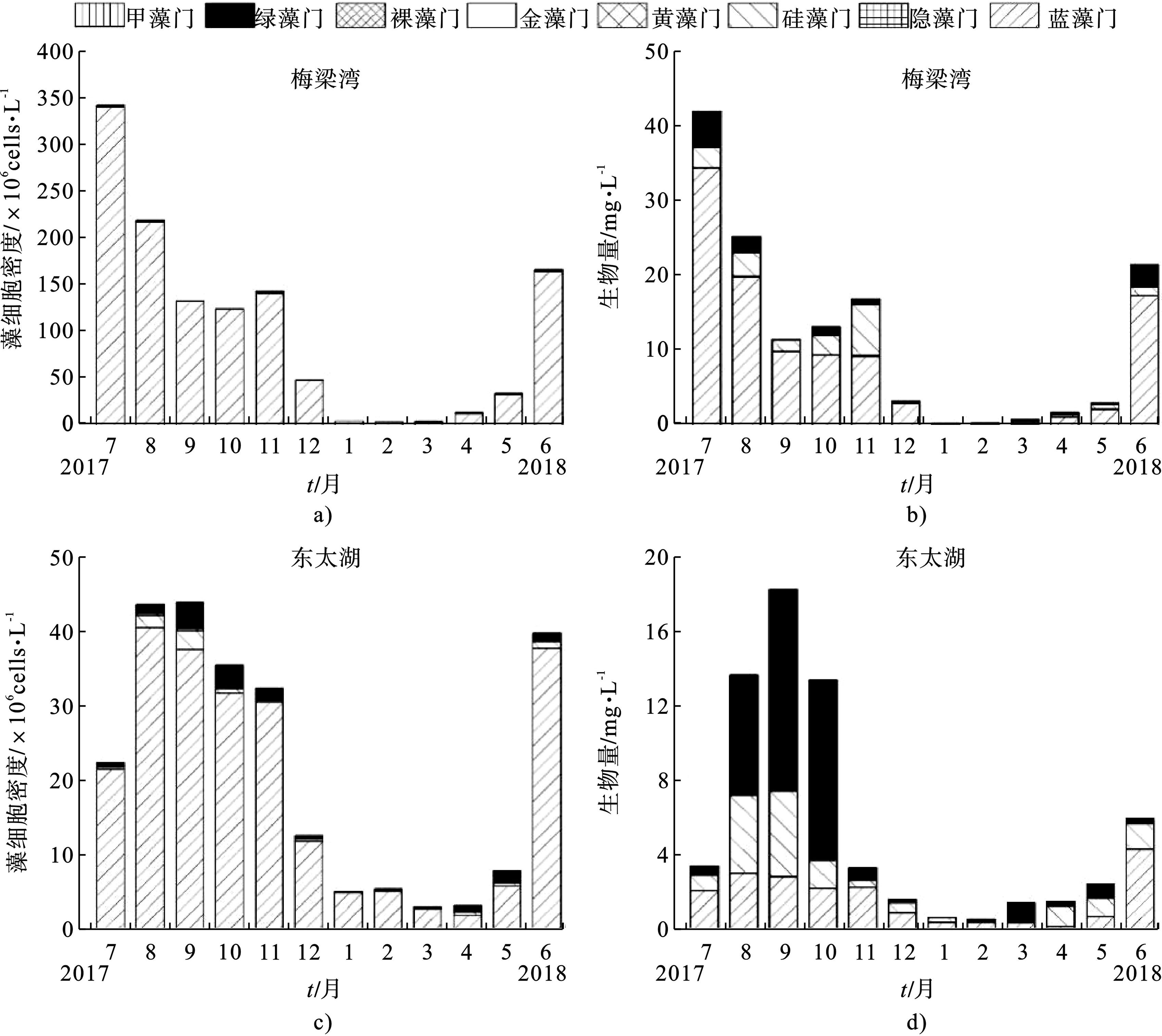
梅梁湾浮游植物数量和生物量具有明显的季节变化,见图4a~b,冬季最低,夏季最高。梅梁湾浮游植物藻细胞密度和生物量在7月达到最大值,分别为343.684×106 cells/L和42.158 mg/L;1月最小,分别为5.185×106 cells/L和0.486 mg/L。调查期间,梅梁湾浮游植物数量全年都以蓝藻门占优势,浮游植物生物量也以蓝藻占优势。
东太湖浮游植物数量和生物量同样具有季节变化,见图4c和图4d,即冬季最低,夏季最高。与梅梁湾不同,东太湖浮游植物细胞密度和生物量最大值出现在9月,分别为43.878×106 cells/L和18.236 mg/L;最小值则分别在3月和2月,分别为3.056×106 cells/L和0.507 mg/L,见图4c~d。调查期间,东太湖与梅梁湾类似,浮游植物数量全年都以蓝藻门占优势,见图4c。与梅梁湾不同的是,2017年8~10月与2018年3~5月,东太湖浮游植物生物量以绿藻或者硅藻占主导地位,见图4d。
-
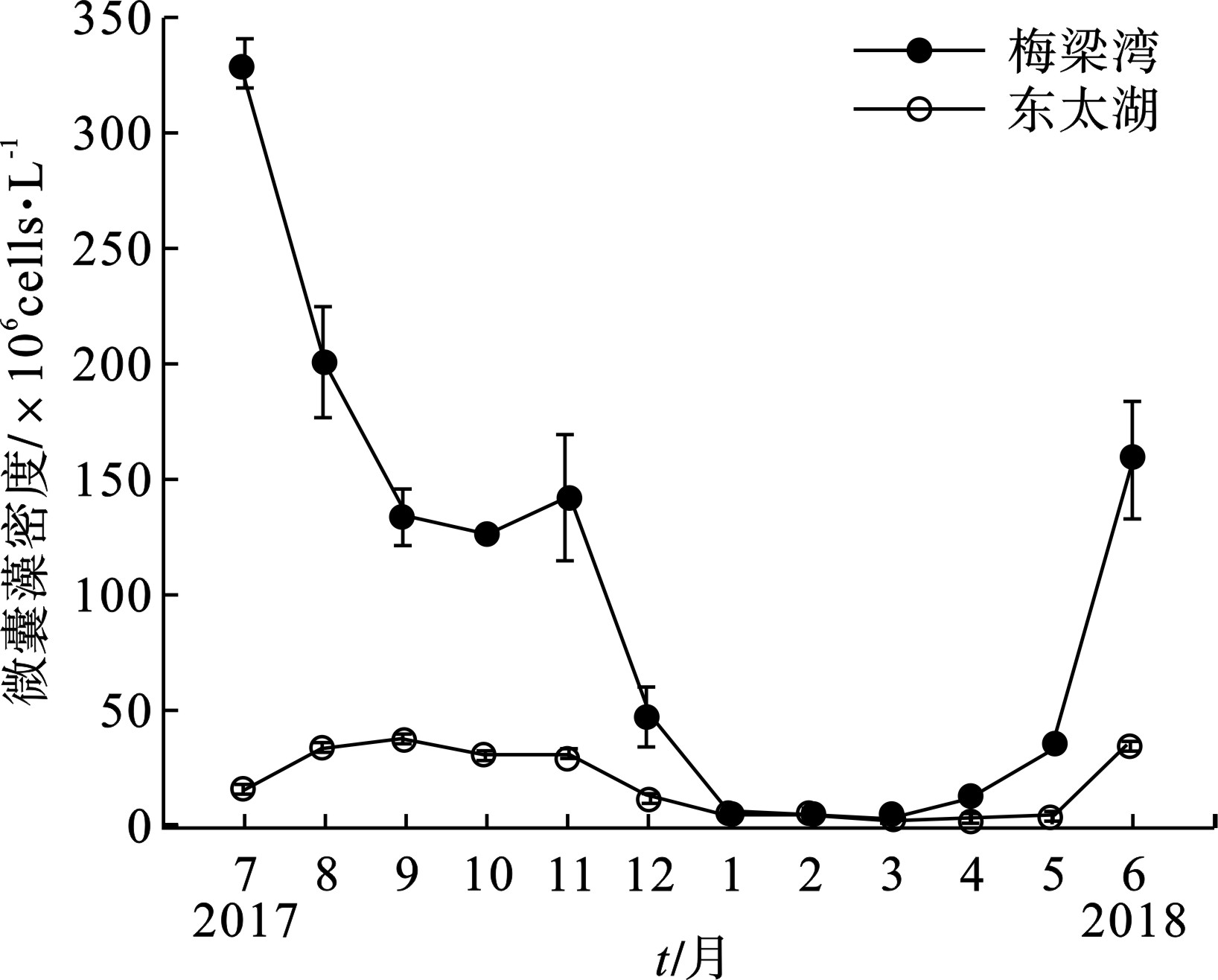
从调查期间的整体情况可看出,微囊藻是梅梁湾和东太湖的优势种,见图5。
图5可知,梅梁湾2017年9月微囊藻占比最高,为99.52%;2017年3月最低,为68.52%。东太湖浮游植物占比较梅梁湾占比低,最高为94.01%,4月微囊藻百分比最低,为57.70%。
除了1~4月外,其他月份梅梁湾微囊藻密度远高于东太湖,梅梁湾7月微囊藻密度最高330.873×106 cells/L,3月最低4.146×106 cells/L;2017年9月东太湖微囊藻密度最高37.488×106 cells/L,4月最低1.844×106 cells/L。梅梁湾湖区微囊藻年均密度(100.12×106 cells/L)显著高于东太湖(17.87×106 cells/L)。因此,梅梁湾湖区比东太湖微囊藻水华更为严重,见图6。
-
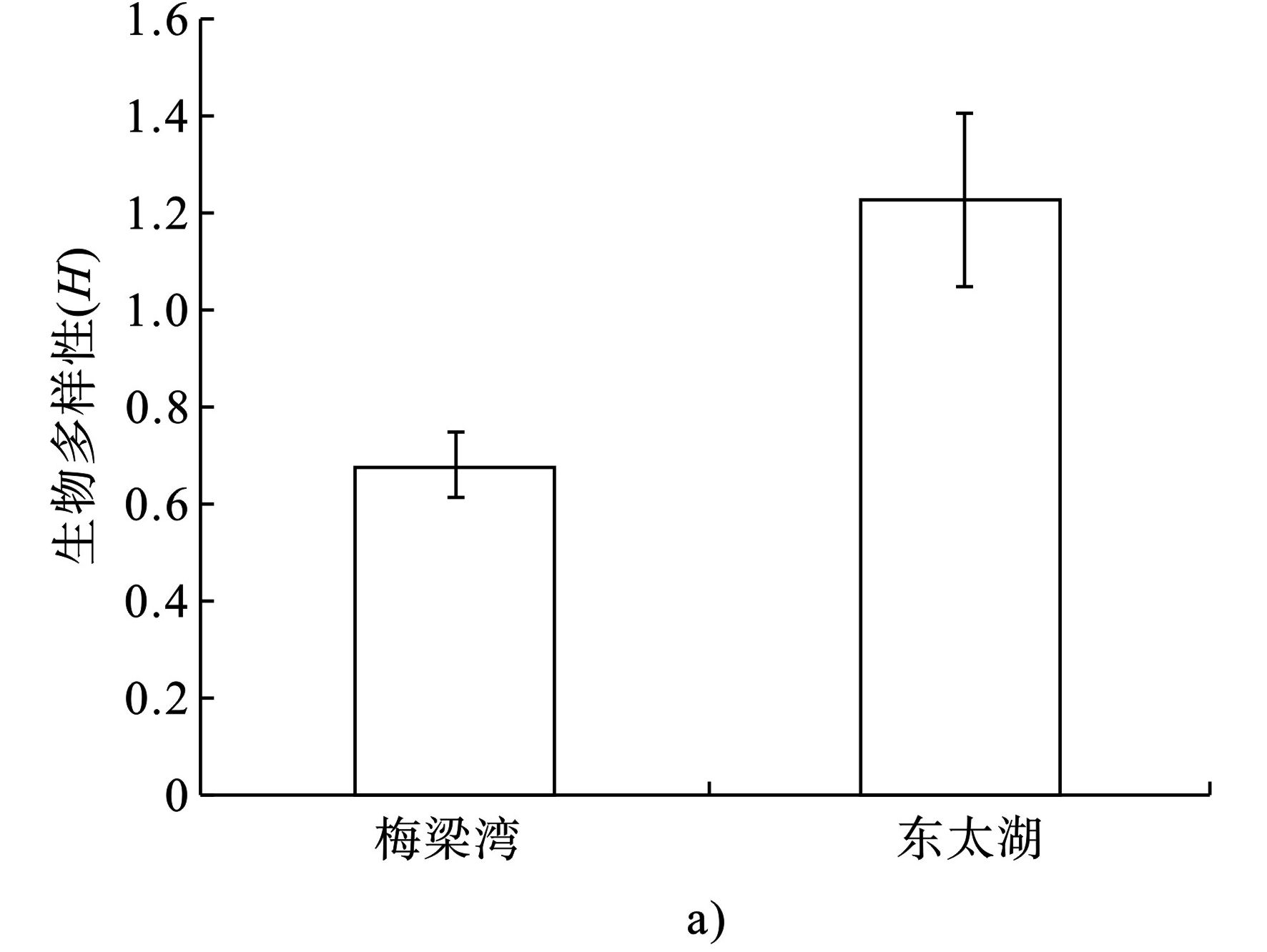
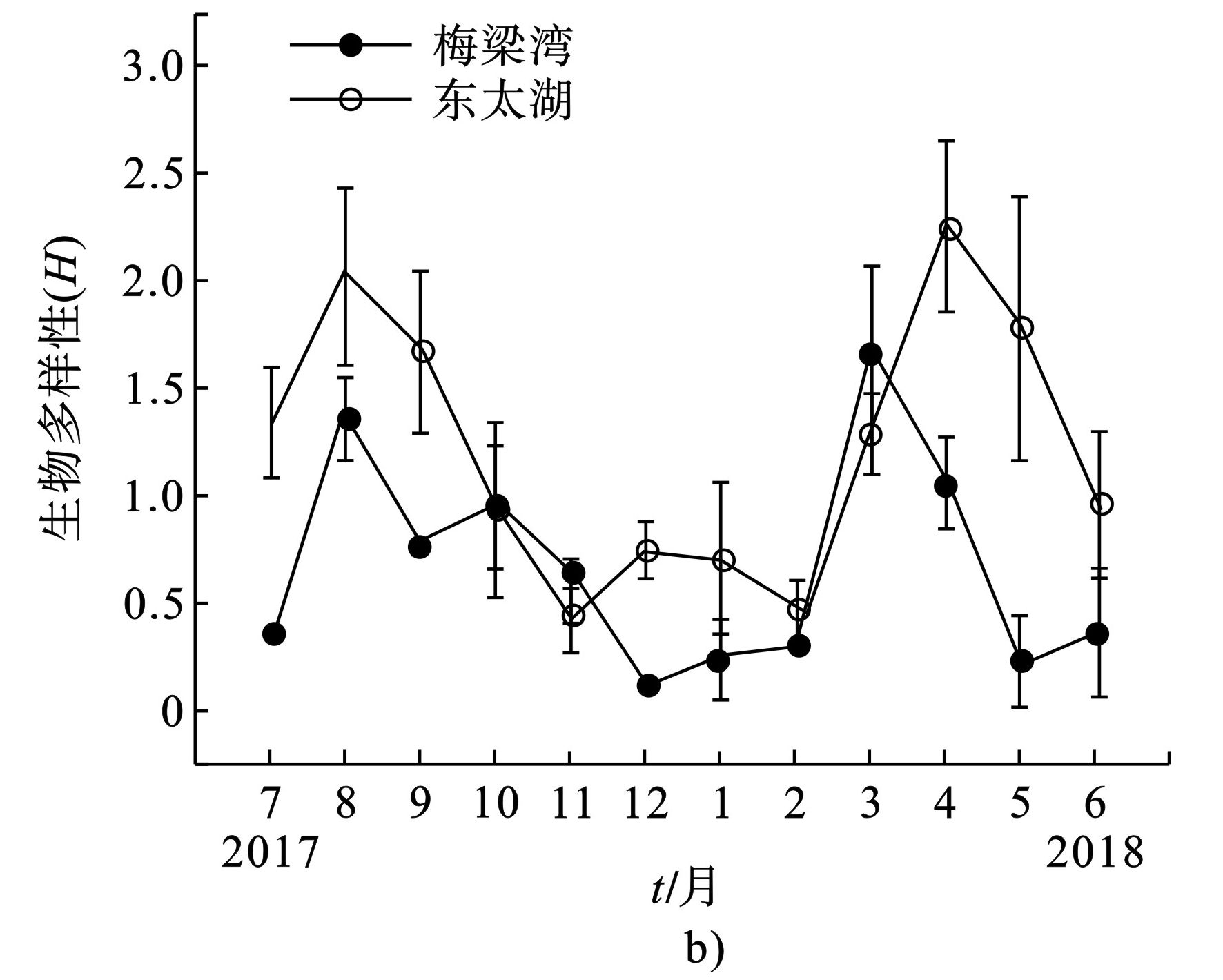
东太湖浮游植物生物多样性指数(H)显著高于梅梁湾(P<0.05),平均值分别为1.22和0.67。两者的生物多样性指数年变化趋势相似,其中东太湖浮游植物生物多样性指数在4月达到最大值,为2.24,其次是8月,为2.03,11月最低,为0.44。梅梁湾在3月H值最大为1.67,其次是8月,为1.03,12月最低,为0.11,见图7。
-
对梅梁湾、东太湖浮游植物数量和生物量与环境因子进行了Pearson相关分析,见表2。
其中,梅梁湾浮游植物数量和生物量与水温、氨氮、硝氮和总磷极显著相关。与梅梁湾不同,东太湖浮游植物数量与水温、总磷显著相关。该研究结果表明,同一湖泊不同湖区影响浮游植物数量和生物量的环境因素不同。
-
环境因子如水温、氮磷营养盐、水动力、浮游动物和水生植物等都是影响浮游植物群落结构的重要因素[14-15]。Pearson相关分析显示,梅梁湾与东太湖湖区浮游植物数量都与水温、总磷显著正相关,见表2。梅梁湾与东太湖同处一个湖泊的不同湖区,水温并没有显著差异,见表1,而梅梁湾总氮和总磷浓度显著高于东太湖(P<0.05);另外,溶解性总氮和溶解性总磷浓度也表现为梅梁湾湖区显著高于东太湖湖区(P<0.05)。该研究表明,梅梁湾湖区可被浮游植物直接利用的氮磷浓度显著高于东太湖,这可能是梅梁湾浮游植物生物量显著高于东太湖的重要原因之一。其次,梅梁湾处于太湖的北部,由于太湖夏季盛行东南风,在风力的作用下,蓝藻容易水平漂移,然后聚集在梅梁湾湖区,所以风力作用是夏季梅梁湾浮游植物生物量显著高于东太湖的原因之一。另外,东太湖的水生植物与藻类竞争氮磷、微量元素和光照[16-17];再者,东太湖的大量水生植物产生的化感物种会抑制藻类的生长和光合作用[18]。因此,氮磷浓度的差异、风力作用和水生植物的竞争与抑制作用可能是东太湖浮游植物数量和生物量显著低于梅梁湾的主要原因。
-
太湖是一个扰动频繁的湖泊,风浪扰动引起底泥悬浮,导致水体透明度下降,进而引起水下光照不足[19],风浪扰动促使微囊藻形成大群体,群体微囊藻比单细胞具有更快的上浮速度,这更有利微囊藻上浮至水体表面获得光照,进而在与其他藻类的竞争中获得优势[20-21]。微囊藻形成群体不利浮游动物摄食,又进一步有利于微囊藻成为优势种[22]。东太湖位于太湖的东部,生长着不同种类的水草,是一个典型的草型湖区。然而,赵凯研究发现持续的高水位和人工水草收割导致2015年太湖水生植被的严重退化,相对于太湖2010年水生植物面积为296.24 km2,2015年太湖水生植物面积减少了52.4 %,缩小为141.15 km2[23]。赵凯[10]研究发现,2015年夏季太湖水生植被分布区面积仅占2014年夏季的20.14 %,2015年冬季水生植被分布区面积仅占2014年冬季的8.94%。水生植物的大面积减少将严重削弱其对浮游植物尤其是微囊藻的竞争和抑制作用。水体富营养以及水生植物减少可能是东太湖微囊藻成为优势种的原因。
杨桂军[24]对太湖梅梁湾调查发现,2007年1~3月以绿藻和硅藻为优势,其他月份则是微囊藻为优势。孟顺龙等[25]对梅梁湾周年调查也发现梅梁湾2008年1~3月主要以绿藻为优势,其他月份以蓝藻中的微囊藻为优势种。本研究中太湖梅梁湾周年调查中所有月份都以微囊藻为优势,微囊藻数量占总浮游植物数量比例范围为68.52%~99.52%。杨桂军[24]报道2006年7月~2007年6月梅梁湾微囊藻年平均密度为0.11×108 cells/L,而本研究梅梁湾微囊藻年平均密度为1.00×108 cells/L,10年后微囊藻密度为10年前的10倍左右。王华等[26]报道从2010~2017年梅梁湾的蓝藻数量呈现逐年增长的趋势,该研究结果表明梅梁湾微囊藻在出现时间和数量方面都呈现扩张趋势。
东太湖浮游植物在20世纪60年代夏季以硅藻为优势种群,数量占总藻类的72%[27]。20世纪50~60年代中期,太湖的蓝藻水华只是偶见于五里湖和鼋头渚附近水域,20世纪70年代中期,大片蓝藻水华被发现于焦山附近,20世纪80年代,五里湖和梅梁湾约有2/5~3/5水域每年夏季出现水华,20世纪90年代中期,整个梅梁湾水域几乎被蓝藻水华覆盖,同时蓝藻水华出现在堂山湾、西部湖区和北部湖心区[27]。2010 年以后,蓝藻水华已扩张到太湖的湖心和东部湖区。在空间分布上,太湖蓝藻水华的扩张沿着梅梁湾和堂山湾逐渐扩展到西部湖区和湖心区,并最终发展至东部湖区[28]。王华等[26]报道从2010~2016年东太湖的蓝藻数量变化不大,而2017年蓝藻数量显著增加,2010年为4.46×106 cells/L,2017年增长为26.51×106 cells/L,2017年是2010年的5.94倍。本研究报道2017年7月至2018年6月东太湖微囊藻年平均密度为17.87×106 cells/L,与文献[26]报道的蓝藻密度比较相近。
-
通过2017年7月至2018年6月对太湖梅梁湾和东太湖浮游植物群落结构和理化因子的调查结论如下。
(1)梅梁湾氮磷浓度显著高于东太湖,梅梁湾浮游植物年平均数量是东太湖4.94倍,梅梁湾浮游植物年平均生物量是东太湖的2.16倍。
(2)梅梁湾微囊藻数量占比为68.52%~99.52%,东太湖微囊藻数量占比为57.70%~94.01%。
(3)太湖蓝藻水华正逐渐扩张至全太湖。
浮游植物群落结构季节变化研究
——以太湖梅梁湾和东太湖为例Study on seasonal variations of community structure of phytoplankton ——Case study of Meiliang Bay and East Taihu Lake in Lake Taihu
-
摘要: 为了解太湖浮游植物群落结构的特征,2017年7月至2018年6月对太湖梅梁湾(藻型湖区)和东太湖(草型湖区)浮游植物群落结构和理化因子进行了调查。结果发现:梅梁湾氮磷浓度显著高于东太湖,梅梁湾浮游植物年平均数量是东太湖4.94倍,梅梁湾浮游植物年平均生物量是东太湖的2.16倍;梅梁湾微囊藻数量占比为68.52%~99.52%,东太湖微囊藻数量占比为57.70%~94.01%。结果表明,太湖草型湖区(如东太湖)正逐渐向藻型湖区(如梅梁湾)发展,研究结果将为太湖管理提供科学依据。Abstract: In order to understand the characteristics of phytoplankton community structure in Lake Taihu, the phytoplankton community structure and physical and chemical factors in Meiliang Bay (algae-dominated lake area) and East Taihu Lake (macrophytes-dominated lake area) were investigated from Jul. 2017 to Jun. 2018. The investigation results found that the concentrations of the nitrogen and phosphorus in Meiliang Bay were significantly higher than these in East Taihu Lake. The annual average abundance and biomass of phytoplankton in Meiliang Bay were 4.94 and 2.16 times of that in East Taihu Lake. The proportions of Microcystis to total algae abundance were 68.52%~99.52% and 57.70%~94.01% in Meiliang Bay and East Taihu Lake, respectively. The investigation results suggested that the macrophytes-dominated lake area (e.g. East Taihu Lake) was gradually developing towards the algae-dominated lake area (e.g. Meiliang Bay) in Lake Taihu, which provided a scientific basis for the management of Lake Taihu.
-
Key words:
- Meiliang Bay /
- East Taihu Lake /
- phytoplankton /
- seasonal variation
-

-
表 1 梅梁湾、东太湖调查期间环境参数均值
理化参数 平均值±标准误差(范围) P值 梅梁湾 东太湖 温度/℃ 17.7±0.1(3.0~25.1) 18.1±0.1(3.0~27.5) 0.733 透明度/cm 34.4±3.2(15.0~86.7) 27.2±3.1(19~33.3) 0.357 pH 7.8±0.1(7.3~8.6) 7.7±0.1(7.3~8.2) 0.568 总氮/mg·L−1 2.729±0.199(1.988~3.200) 2.027±0.208(1.523~2.827) <0.001 溶解性总氮/mg·L−1 1.645±0.231(1.024~2.945) 1.225±0.101(0.782~2.117) 0.023 硝氮/mg·L−1 0.806±0.118(0.350~1.790) 0.584±0.054(0.181~1.354) 0.147 氨氮/mg·L−1 0.382±0.060(0.290~0.538) 0.294±0.027(0.190~0.407) 0.077 总磷/mg·L−1 0.224±0.017(0.052~0.270) 0.118±0.005(0.023~0.203) 0.027 溶解性总磷/mg·L−1 0.062±0.004(0.026~0.117) 0.030±0.004(0.012~0.037) 0.004 正磷酸盐/mg·L−1 0.027±0.003(0.005~0.056) 0.021±0.003(0.005~0.033) 0.243 悬浮颗粒物/mg·L−1 36.22±1.98(13.67~75.33) 37.17±5.11(12.66~72.00) 0.895 COD/mg·L−1 20.79±2.26(11.13~42.27) 18.00±1.02(10.22~35.68) 0.483 底泥总氮/mg·L−1 0.886±0.066(0.576~1.777) 0.944±0.080(0.560~1.679) 0.751 底泥总磷/mg·g−1 0.154±0.006(0.084~0.247) 0.152±0.006(0.098~0.236) 0.919 叶绿素a/μg·L−1 57.10±3.10(7.77~150.58) 23.19±0.68(10.11~50.56) 0.025  下载: 导出CSV
下载: 导出CSV
表 2 梅梁湾、东太湖浮游植物数量和生物量与环境因子相关性分析
环境参数 梅梁湾 东太湖 数量 生物量 数量 生物量 水温 0.854** 0.828** 0.584* 0.544 pH −0.138 −0.187 0.29 0.411 透明度 −0.48 −0.436 0.531 0.311 总氮 0.353 0.375 −0.188 −0.2 溶解性总氮 −0.439 −0.423 −0.338 −0.04 氨氮 −0.810** −0.763** 0.036 0.25 硝氮 −0.752** −0.703* −0.442 −0.238 总磷 0.963** 0.940** 0.640* 0.32 溶解性总磷 0.494 0.481 0.378 0.41 悬浮颗粒物 0.521 0.434 −0.402 −0.099 注:*表示P<0.05,**表示P<0.01。
下载: 导出CSV
-
[1] HORNE A J, GOLDMAN C R. Limnology[M]. (2nd ed. ). New York: Mc Graw-Hill Companies, 1994: 226-264. [2] HÅKANSON L, BOULION V V. A general dynamic model to predict biomass and production of phytoplankton in lakes[J]. Ecological Modelling, 2003, 165(2−3): 285 − 301. doi: 10.1016/S0304-3800(03)00096-6 [3] GAMEIRO C, CARTAXANA P, BROTAS V. Environmental drivers of phytoplankton distribution and composition in Tagus Estuary, Portugal[J]. Estuarine, Coastal and Shelf Science, 2007, 75(1−2): 21 − 34. doi: 10.1016/j.ecss.2007.05.014 [4] THIÁHAUT G, TIXIER G, GUEROLD F, et al. Comparison of different biological indices for the assessment of river quality: Application to the upper river Moselle (France)[J]. Hydrobiologia, 2006, 570(1): 159 − 164. doi: 10.1007/s10750-006-0176-2 [5] 窦鸿身, 姜加虎. 中国五大淡水湖[M]. 北京: 中国科学技术出版社, 2003. [6] CHEN Y, QIN B, TEUBNER K, et al. Long-term dynamics of phytoplankton assemblages: Microcystis-domination in Lake Taihu, a large shallow lake in China[J]. Journal of Plankton Research, 2003, 25(4): 445 − 453. doi: 10.1093/plankt/25.4.445 [7] QIN B Q, ZHU G W, GAO G, et al. A drinking water crisis in Lake Taihu, China: linkage to climatic variability and lake management[J]. Environmental Management, 2010, 45: 105 − 112. [8] 水利部太湖流域管理局, 江苏省水利厅, 浙江省水利厅, 等. 太湖健康报告[R/OL]. [2021-12-28] http://www.tba.gov.cn/slbthlyglj/thjkzkbg/content/slth1_21eeb1db2bd54415b685221ba98caf07.html. [9] 朱伟, 谈永琴, 王若辰, 等. 太湖典型区2010—2017年间水质变化趋势及异常分析[J]. 湖泊科学, 2018, 30(2): 296 − 305. doi: 10.18307/2018.0202 [10] 赵凯. 太湖水生植被分布格局及演变过程[D]. 南京: 南京师范大学, 2017. [11] 金相灿, 屠清瑛. 湖泊富营养化调查规范[M]. 北京: 中国环境科学出版社, 1990. [12] 胡鸿钧, 魏印心. 中国淡水藻类: 系统分类及生态[M]. 北京: 科学出版社, 2006. [13] 章宗涉, 黄祥飞. 淡水浮游生物研究方法[M]. 北京: 科学出版社, 1991. [14] ARHONDITSIS G B, WINDER M, BRETT M T, et al. Patterns and mechanisms of phytoplankton variability in Lake Washington (USA)[J]. Water Research, 2004, 38(18): 4013 − 4027. doi: 10.1016/j.watres.2004.06.030 [15] HANEY J F, HALL D J. Sugar-coated Daphnia: a preservation technique for cladocera[J]. Limnology and Oceanography, 1973(18): 331 − 333. [16] ENGELHARDT K A M, RITCHIE M E. Effects of macrophyte species richness on wetland ecosystem functioning and services[J]. Nature, 2001, 411: 687 − 689. doi: 10.1038/35079573 [17] 戴莽, 倪乐意, 谢平, 等. 利用大型围隔研究沉水植被对水体富营养化的影响[J]. 水生生物学报, 1999, 23(2): 97 − 101. doi: 10.3321/j.issn:1000-3207.1999.02.001 [18] SØNDERGAARD M, MOSS B. Impact of submerged macrophytes on phytoplankton in shallow freshwater lakes [M]. In: Jeppesen E, Søndergaard M, Søndergaard M &Christoffersen K(Eds), The Structuring Role of Submerged Macrophytes in Lakes. New York: Springer, 1998: 115-132. [19] TANG X M, KRAUSFELDT L E, SHAO K, et al. Seasonal gene expression and the ecophysiological implications of toxin Microcystis aeruginosa blooms in Lake Taihu[J]. Environmental Science & Technology, 2018, 52: 11049 − 11059. [20] 秦伯强, 杨桂军, 马健荣, 等. 太湖蓝藻水华“暴发”的动态特征及其机制[J]. 科学通报, 2016, 61(7): 759 − 770. [21] QIN B Q, YANG G J, MA J R, et al. Spatiotemporal changes of cyanobacterial bloom in large shallow eutrophic Lake Taihu, China[J]. Frontiers in Microbiology, 2018, 9: 451. doi: 10.3389/fmicb.2018.00451 [22] OLIVER R L, GANF G G. Freshwater blooms[M]. Dordrecht: Kluwer Academic Publishers, 2000: 149-194. [23] 许雪敏. 太湖水生植物遥感监测及演变驱动因素分析[D]. 西安: 西安科技大学, 2018. [24] 杨桂军. 浮游植物对浮游动物和营养盐胁迫的响应研究[D]. 南京: 中国科学院南京地理与湖泊研究所, 2008. [25] 孟顺龙, 陈家长, 胡庚东, 等. 2008年太湖梅梁湾浮游植物群落周年变化[J]. 湖泊科学, 2010, 22(4): 577 − 584. [26] 王华, 陈华鑫, 徐兆安, 等. 2010--2017年太湖总磷浓度变化趋势分析及成因探讨[J]. 湖泊科学, 2019, 31(4): 919 − 929. doi: 10.18307/2019.0421 [27] 孙顺才, 黄倚平. 太湖[M]. 北京: 海洋出版社, 1993. [28] 张民, 阳振, 史小丽. 太湖蓝藻水华的扩张与驱动因素[J]. 湖泊科学, 2019, 31(2): 336 − 344. doi: 10.18307/2019.0203 -
 点击查看大图
点击查看大图
图( 9) 表( 2)
计量
- 文章访问数: 4038
- HTML全文浏览数: 4038
- PDF下载数: 28
- 施引文献: 0






