

-
城市内河是城市废弃物、生活污水和暴雨径流的主要汇集地,也是城市水循环的重要输送环节,被喻为“城市动脉”,起到调节城市微气候、美化城市环境等关键作用。我国城镇化进程加速了城市规模的扩张,随之而来的是城市内河水体污染问题日益凸显,内河生态系统濒临崩溃,水体黑臭是城市内河最易发生的现象[1]。自2015年起,我国开启了城市黑臭水体的摸排与大力整治,形成了“控源截污、内源治理、生态修复、活水提质、长制久清”的治理模式。到了2020年,城市内河水体的黑臭现象得到了根本的改善,但返黑返臭现象时有发生。究其根源,是城市内河受到周边的外源排放负荷冲击,超出了生态系统的调节界限[2]。
尽管人们对于水环境保护的认识日益提高,并且正在努力保护城市河流资源的可持续发展,但城市的高速发展不可避免地导致河流环境的恶化加剧[3]。由于高负荷污水排放和水体微生物的特性(特别是多样性和群落结构)可能会受到物理化学和生物参数的空间变异性的影响,这些参数可作为环境条件的指标[4]。以往大多数评估城市人类活动对地表水影响的实验室和现场研究都集中在物理化学指标上,如重金属[5]、碳氢化合物[6]和藻类特征[7]。对城市内河水体中微生物群落多样性的深入了解仍十分匮乏,并且相较于海洋或湖泊生态系统的研究更少。
微生物群落作为环境胁迫的敏感指标,既是生态系统的初级生产者和主要分解者,又是物质循环中的重要成员与物质和能量的贮存者,对生态系统的稳定起决定性作用,同时能够快速反映河流生态系统的状态,菌群的空间分布特征是与环境因子综合作用的结果。由于内河水体的补给以大气降雨、径流汇入、沿河点源排放为主,补给源的水质对于内河的微生物群落组成具有决定性作用[8]。研究环境因子对生物群落多样性、结构、组成以及空间分布格局是生态学的重要内容之一[9]。在城市内河水体的环境因子中,水体理化指标能够直接影响微生物群落的空间分布特征。对黄河中的浮游细菌群落主要受到pH、溶解有机碳(DOC)的影响 [10],我国华南地区的东江中氨氮(NH3-N)和硝态氮(NO3-N)是影响河流浮游细菌群落的主要环境因子[11]。瑞典的14个湖泊中氮磷营养盐(TN、TP)是影响微生物群落多样性的主要环境因子[12]。因此,了解城市内河中微生物群落的空间分布和微生物多样性对于监测生态系统的健康和功能至关重要[13]。
不同于传统的微生物研究手段局限于可培养微生物的研究,近年来高速发展的高通量测序技术,能够快速、系统地分析环境样品中的微生物组成、结构、多样性以及认识微生物的生态功能[14]。这极大地促进了微生物生态学的发展,也使得高通量测序技术能广泛地应用到研究各种环境介质的微生物中[15]。众多学者针对不同类型水体,如海洋[16]、湖泊[17]、天然河流[18]、城市内河[19]等高通量测序技术进行了关于微生物的赋存状态、微生物与自然环境因子之间的响应关系、微生物与其他污染物的交互影响机制、在外来污染源如工业污水、生活污水等影响下微生物群落的响应特征等研究。尤其针对城市内河,研究主要集中在城市内河水体中微生物的空间分布、城市内河水体的群落结构和多样性、城市内河水体生物群落结构受环境因素的影响、城市黑臭水体成因和治理措施等[20],但是,目前对易返黑返臭时期的城市内河水体的微生物的群落的研究相对较少。
本项目以夏末秋初芜湖城区典型城市内河段作为研究对象,通过16S rRNA高通量测序技术研究了3条内河水体微生物群落的结构和多样性,探讨主要环境因子与微生物群落之间的关系,为预防城市内河水体的返黑返臭现象提供理论基础。
-
芜湖地处长江中下游平原,属亚热带季风气候,四季分明,气候多变,年平均降水量为1184.4 mm。地表水和地下水资源丰富,全市水面面积达478 km2,占总面积14.4%。其中,青戈江是芜湖市境内最主要的河流,其支流流经整个戈江区,包括了复杂的城市内河水体。内河流经居民小区、商业区、工业区等不同功能区,接纳降雨地表径流、管网泄漏以及部分点源汇入,水质呈现季节性波动。论文选取的3条典型城市内河河段,中央城(ZYC)河段、汇成(HC)河段和中山南路(ZSNL)河段均位于弋江区,中央城河段(ZYC)长约1.5 km ,周边主要有中央城会展中心、驾校、商业区等,汇成河段(HC)长约3.5 km ,流经汇成小区、芜湖市第四人民医院,中山南路(ZSNL)河段长约4.5 km,流经居民区、学校、商业区等,各河段均有多个排污口,中山南路正在修建市政管道。
-
根据污染源类型和排污口分布情况,在3条河段上布设了18个采样断面(图1)。其中,中央城河段布设3个采样点(ZYC1-ZYC3),汇成河段布设6个采样点(HC1-HC6),中山南路河段布设9个采样点(ZSNL1-ZSNL9)。
2020年9月4日、5日,按照《水质采样技术指导》(HJ 494—2009)进行样品采集与现场固定,用于微生物测试的水样采集后放入-4 °C恒温箱中暂存,委托上海派森诺生物科技有限公司采用 Illumina MiSeq高通量测序技术测序。采样断面经纬度如表1所示。
-
采用便携式水质检测仪(T3WS-GP)测试pH、水温(WT)、DO、ORP 、EC等参数,水体的透明度采用塞氏盘法。 COD、 BOD5、 TN、 TP、 NH3-N和 Chla的质量浓度按国标法进行测定。
-
委托上海派森诺生物科技有限公司采用Illumina MiSeq高通量测序技术进行微生物测序,采用MoBio/QIAGEN公司的DNeasy PowerWater Kit对3条城市内河水体中的微生物总DNA基因组进行提取,采用分子荧光分光光度计(Quantifluor-ST fluorometer,Promega,E6090;Quant-iT PicoGreen dsDNA Assay Kit,Invitrogen,P7589),在260 nm和280 nm处分别测定DNA的吸光值,检测DNA的浓度,并用1%的琼脂糖凝胶电泳检测DNA的质量。调整DNA溶液浓度,DNA工作液保存于4 ℃,储存液保存于−20 ℃。高通量测序采用引物序列为正向引物338F(5′-ACTCCTACGGGAGGCAGCA-3′)和反向引物806R(5′-GGACTACHVGGGTWTCTAAT-3′)[21]扩增城市内河水体微生物细菌的16S rRNA基因V3-V4区域。PCR 扩增首先对16S rRNA基因可变区进行扩增,纯化,Bio Tek酶标仪检测。最终得到每个扩增序列变体 (amplicon sequence variants,ASVs)代表序列的分类学信息。
高通量测序结果显示,9个水样共得到384273条高质量基因序列, 以100%相似度划分,得到11732条不同的ASVs,单个水样的 ASV数量在1587到3662之间,平均值为2176。优质序列的序列长度分布在 405—431 bp,平均值为410 bp。3条河流浮游细菌群落共66门,969属。
-
基于EXCEL处理的原始实验数据,采用SPSS.19.0软件计算Spearman秩相关系数,确定环境因素与细菌群落的多样性或丰度之间的关系,并进行单因素方差分析(one-way ANOVA)检验不同组别环境因子的差异。采用QIIME2软件处理微生物原始数据,DADA2质控后产生的每个去重的ASVs特征序列,ggplot2软件包计算Alpha多样性指数等。β多样性则使用vegan进行PCoA分析,使用R基本软件包进行了蒙特卡罗置换测试即冗余判别分析(RDA)以揭示细菌组合与环境因素之间的关系,显示了与细菌群落显著相关的环境因子。
-
采用单因素方差分析(one-way ANOVA)方法分析芜湖城市内河水质(表2)。各项水质指标大体上呈现为中央城河段<汇成河段<中山南路河段。3条河段的温度在26.8—29.6 ℃之间, pH值变化范围为7.57—9.49,平均溶解氧量表现为中央城河段(2.7 mg·L−1)<汇成河段(5.91 mg·L−1)<中山南路河段(9.89 mg·L−1)。单因素方差分析(one-way ANOVA)显示,3条河段的TN、TP、COD 、NH3-N差异显著。TN含量均高于(GB3838-2002)中V类水标准值2 mg·L−1。汇成河段COD和TN含量在HC4处达到最高,分别为19.9 mg·L−1和5.93 mg·L−1,而Chl a含量在HC4处最低为30 mg·L−1,该断面处有芜湖市第四人民医院和汇成小区污水排放口,导致该处TN和NH3-N含量超标。中央城河段的COD、BOD5、NH3-N、TP在断面ZYC2达到最大值分别为12.7、8.59、1.7、0.153 mg·L−1,其中,COD、BOD5、NH3-N超过了《地表水环境质量标准》(GB3838-2002)Ⅳ类水标准值。中山南路河段COD含量在ZSNL5处最大为31.5 mg·L−1, BOD5含量最低仅为3.50 mg·L−1, NH3-N和Chl a含量最高分别为8.64 mg·L−1和157 mg·L−1。该断面处的污水管道在维修,有黑褐色污水流入河段,根据城市黑臭水体分级标准(表3),采样点ZYC2和ZSNL5水体达到轻度黑臭,表明中央城河段和中山南路河段已出现返黑返臭现象。
-
由表4 可知,各采样点的覆盖度Goods_coverage指数均接近1,说明本次测序深度基本覆盖了样本中的所有物种。Shannon指数高且差异不大,说明三条城市内河的生物多样性高。各河段的采样点的Simpson指数低,说明三条城市内河的物种丰富度高,即生物多样性高,这一结论与Shannon指数所得结果一致。各采样点的均匀度Pielou’s PD指数总体上较高,且各菌落之间差异性不大。Observed species指数显示三条城市内河河段的各采样点的菌落中含有的物种数目的平均值:汇成河段>中央城河段>中山南路河段。与其他河段的采样点相比汇成河段的采样点HC-1的Chao1 指数大,说明菌落的物种丰富度高,这可能与第四人民医院的污水排放有关。中央南路河段的各采样点的Chao 1 指数相对较小,即菌落的物种丰富度相对较低,这可能与正在维修市政管线有关,这与圣安娜河城市段的微生物群落的研究结果一致[22]。另一方面,中央城河段和中山南路河段的菌落的多样性和ASV丰富度低于汇成河段,这可能与所受纳污水的种类和数量有关[23]。但是3条城市内河河段的采样点的Chao 1指数都大于ASV数量,表明样品中的细菌存在未测明序列有待进一步探索。
通过alpha多样性分析,可以较清晰地比较不同河段各断面的菌落多样性,反映生境内微生物的丰富度和均匀度(图2)。
各采样点的Chao1指数和Shannon指数均存在明显的空间变化特征,汇成河段的Chao1指数和Shannon指数明显大于中央城河段和中山南路河段,不同采样点的多样性指数(Chao1指数、Shannon指数)的变化与ASV变化一致,相互印证。Shannon指数最大值分别为8.85、9.10、8.36,即汇成河段的微生物群落多样性优于中央城河段,且中央城河段的微生物群落多样性高于中山南路河段。
-
采用主坐标分析(Principal coordinates analysis,PCoA),探究芜湖城市内河水体细菌群落结构差异(图3),各采样点在二维图中的距离可以反映群落结构的相似程度,样点之间距离越近表示群落结构越相似。3条河段样点的组内距离较近而组间距离较远,说明受到不同污染的城市内河水体的微生物群落结构存在明显的空间差异性。除中山南路河段外,同一河段的各采样点间具有相似的微生物群落结构,而3条河段间的微生物群落差异较大。这可能与同河段的各采样点与污染物排放口的距离,河段的水体流速以及是否有其他外源输入有一定关联。
-
3条河段的细菌群落在门分类水平上组成如图4所示,其中细菌相对丰度小于1%的物种和未知物种归为Others。研究区的浮游细菌群落结构在门水平上具有较高的多样性.主要包括变形菌门(Proteobacteria)、放线菌门(Actinobacteria)、蓝细菌门(Cyanobacteria)、拟杆菌门(Bacteroidetes)、疣微菌门(Verrucomicrobia)和厚壁菌门(Firmicutes)。中央城河段变形菌门相对丰度最低,约占29.29%—33.46%,放线菌门相对丰度最高,约占39.25%—43.48%,蓝细菌门(Cyanobacteria)约占10.45%—12.40%,拟杆菌门(Bacteroidetes),约占9.07%—10.48%,疣微菌门(Verrucomicrobia)约占1.25%—1.64%;汇成河段变形菌门约、占33.43%—38.36%,放线菌门约占29.01%—34.43%,蓝细菌门(Cyanobacteria)相对丰度最高,约占17.91%—22.02%,拟杆菌门(Bacteroidetes)约占5.92%—8.88%,疣微菌门(Verrucomicrobia)约占0.5%—3.32%;中山南路河段变形菌门相对丰度最高,约占42.04%—59.84%,放线菌门相对丰度约占13.44%—40.42%,蓝细菌门(Cyanobacteria)约占1.52%—14.32,拟杆菌门(Bacteroidetes)约占8.69%—11.55%,疣微菌门(Verrucomicrobia),约占0.25%—3.32%。内河水体的变形菌门(Proteobacteria)主要包括α-变形菌纲(α-Proteobacteria)、β-变形菌纲(β-Proteobacteria)和γ-变形菌纲(γ-Proteobacteria)。3条河流都检测出大量的蓝藻细菌门(Cyanobacteria),汇成河段的疣微菌门(Verrucomicrobia)明显较高,另两条河流疣微菌门(Verrucomicrobia)的相对丰度则相对较小。
3条河段的优势菌门主要包括:变形菌门(Proteobacteria)、放线菌门(Actinobacteria)、蓝细菌门(Cyanobacteria)、拟杆菌门(Bacteroidetes),这与澜沧江流域和珠江下游细菌群落的优势菌门十分相似[22]。与中央城河段和中山南路河段相比,汇成河段水中的蓝细菌门(Cyanobacteria)明显较高,且从上游到下游有显著增加的趋势。有研究报道蓝细菌对调节水体溶解氧的平衡至关重要 [23],这可能与汇成河段未出现返黑返臭迹象有关。
-
3条河段在属分类水平上的浮游细菌群落组成如图5所示,水样中细菌涵盖了969个属,其中相对丰度大于1%包括16个属。中央城河段相对丰度较高的菌属有hgcI_clade、叶绿菌属(Chloroplast)、CL500-29_marine_group、嗜冷杆菌属(Psychrobacter),所占比例范围分别为27.99%—32.83%、9.99%—7.48%、4.78%—5.16%、1.57%—1.62%。汇成河段以hgcI_clade、叶绿菌属(Chloroplast)、CL500-29_marine_group、嗜冷杆菌属(Psychrobacter)为主要优势菌属,所占比例范围分别为17.42%—21.28%、12.05%—16.81%、3.90%—5.37%和2.21%—2.89%。中山南路河段则主要是hgcI_clade、叶绿菌属(Chloroplast)、CL500-29_marine_group、嗜冷杆菌属(Psychrobacter),所占比例范围分别为6.07%—27.88%、1.47%—12.10%、1.29%—3.65%和2.66%—3.76%。
虽然3条河段的优势菌门相似,但由于污染类型不同,各采样点的优势菌属的数量以及组内菌属的比例的分布都存在明显的空间差异性。中山南路河段组内差异最大,中央城河段和汇成河段微生物群落分布较为均匀。变形菌门包含了γ-proteobacteria纲、δ-proteobacteria纲和α-proteobacteria纲3类,具有丰富的代谢多样性,对于 C、N、P 等营养元素的循环以及各种污染物的降解具有重要作用[24]。变形杆菌门是优势菌中数量最多的门,在水体中以γ-变形菌为主。γ-变形菌被认为是富营养化湖泊中最丰富的一类细菌[2],水体样品中γ-变形菌丰度的显著增加与TN、TP和NH3-N含量呈正相关,这可能是由生活污水中的高营养源驱动的[25]。值得注意的是,在HC-2、和HC-3样品中存在少量γ-变形菌(γ-proteobacteria)中的不动杆菌属(Acinetobacter)细菌(图5),此处有医院排污口,有研究报道不动杆菌属(Acinetobacter)是一种新兴的医院致病菌[26],医院排放的废水是河流中致病性微生物的主要来源。因此,该属的丰度可作为医院废水排放的敏感指标的参考。拟杆菌门(Bacteroidetes)是城市内河水体和沉积物中的另一个主要门(图4)。该门中的许多细菌与哺乳动物的肠道微生物群落有关[27],该门内的一些物种已被提议作为有效的替代粪便指标[28]。城市水体中拟杆菌的较高丰度可能是由来自居民区的未经处理的废水造成的,这些废水可能含有硝酸盐、氨、粪便等污染物。而属于拟杆菌门(Bacteroidetes)的黄杆菌是淡水中特别常见和丰富的异养生物,在碳循环中发挥关键作用,如藻类有机物或生物聚合物生物降解。研究数据显示,中山南路下游水中黄杆菌(Flavobacterium)丰度较高,与近期有大量有机废物排入相关。厚壁菌门(Firmicutes)是识别人类粪便的重要指标之一[29],在HC-2、HC-3污水出口附近取样的厚壁菌门(Firmicutes)丰度显著较高,这可能与未经处理的粪便污水有关(图4)。汇成河段主要是周围居民的生活污水和医院的医疗废水排放地,其中拟杆菌门(Bacteroidetes)和厚壁菌门(Firmicutes)含量约为中央城河段和中山南路河段的两倍,与已报到的研究结果一致[30-31],表明不同类型的废水排放导致微生物群落发生了变化。微生物群落是由不同的环境因素驱动,这意味着城市内河微生物群落可能进化出系统发育的通用性来应对来自城市的污染物[32]。
-
通过冗余分析确定了芜湖市3条城市内河水体理化指标与细菌群落的相关性(图6)。第一主轴解释了67.56%的优势菌门群落变化,而第二主轴解释了27.20%优势菌门群落变化(图6)。冗余分析显示,总氮和水温在3条河段的水生细菌群落分布中起关键作用。此外,酸碱度、溶解氧、电导率、氨氮、总磷和叶绿素a也在一定程度上影响着水样中细菌群落的组成。优势菌门与环境因子的Pearson相关性分析如表5所示,变形菌门(Proteobacteria)与WT、TP、TN、Chla、NH3-N显著性正相关,放线菌门(Actinobacteria)与WT、TN、Chla、NH3-N显著负相关,蓝细菌门(Cyanobacteria)和疣微菌门(Verrucomicrobia)与DO显著正相关,厚壁菌门(Firmicutes)与TN、Chla显著性正相关,WT、DO、TN、TP、Chla、NH3-N是造成门水平群落结构差异的主要驱动因子,这与RDA结果显示基本一致。
总磷是与地表水微生物群落变化显著相关的最重要的决定因素。磷被普遍认为是流动水体中生物代谢的重要驱动力,需要小心管理,以避免由城市化造成的富营养化影响。总氮含量是衡量水质的重要指标之一,而三个河段的总氮都出现了严重超标[33]。RDA数据显示,总氮浓度对微生物群落组成的变化有显著贡献,表明氮是另一个重要相关因素,这与以前对淡水的研究一致[34]。此外,氮磷化学计量、酸碱度、温度[35]、溶解氧[36]和重金属也会对河流生态系统中微生物群落产生影响[37]。诸多因素相互作用,影响着芜湖市城市内河的微生物群落。
虽然河水中的细菌群落受营养物浓度、重金属污染、温度和流速等多种因素的影响,但是地理环境、采样策略和观测参数的不同,对河水细菌群落影响因素的研究会呈现出不同的结果[38]。一些对城市河流的一次性采样研究发现,河流中的细菌群落呈现出受其他影响因素驱动的趋势。例如在泰晤士河下游,拟杆菌属占优势的群落向非拟杆菌属占优势的群落转变,这主要是由水的停留时间而不是水化学因子驱动的[39],多瑙河研究发现了同样的趋势,但将其归因于下游土壤和地下水对细菌的影响减少[40]。因此,研究城市内河水体微生物群落的主要影响因素,还需要综合水文气象的影响,采取全年多时间段进行研究分析。
-
(1)高通量测序结果表明,3条河段9个水样的高通量测序共得到384273条高质量基因序列,获得浮游细菌群落66门,969属。在门分类水平上,细菌群落中优势门为变形菌门 (Proteobacteria),其次是放线菌门 (Proteobacteria)。在属分类水平上,3条河段细菌群落中相对丰度最高的是hgcI_clade (20.45%),其次为Chloroplast (10.73%)和CL500-29_marine_group (3.99%),且从属分类水平上存在大量未知属细菌。
(2)3条河段细菌多样性丰富,汇成河段水样的Chao1指数和Shannon指数明显高于中央城河段和中山南路河段。其中汇成河段小区污水排放口排放的大量生活污水的排放造成γ-变形菌(γ-Proteobacteria)数量的显著增加,该属的细菌大量繁殖,使水体优势菌群结构单一化,多样性指数下降,再加上外源点源汇入,导致汇成河段的蓝细菌门数量高于另两条河段,这可能与汇成河段未出现返黑返臭迹象有关联。
(3)RDA分析表明,影响浮游细菌群落结构变化的关键环境因子为pH、温度、TN、TP、DO、Chl a、COD和EC。浮游细菌群落结构差异的根本原因是三条河段的空间差异。中央城河段主要受EC的影响,汇成河段主要受EC和DO的影响,中山南路河段主要受TN、TP的影响。
芜湖城市内河水体微生物群落结构特征及其影响因素
Microbial community structure and its influencing factors in urban river water in Wuhu
-
摘要: 为研究夏末秋初芜湖城市内河水质波动原因,在综合考虑河段周边土地利用类型、污染源排放等因素的基础上,论文选择芜湖三条代表性城市内河河段(中央城河段、汇成河段、中山南路河段)作为研究对象,布设了18个监测断面,分析了pH、溶解氧(DO)、电导率(EC)、氨氮(NH3-N)和总磷(TP)以及叶绿素a(Chla)等水质指标,采用16S rRNA高通量测序技术测试了微生物群落,分析了微生物群落结构特征。研究结果表明,芜湖市三条城市内河河段细菌多样性丰富,主要菌门种类相似,包括变形菌门(Proteobacteria)、放线菌门(Actinobacteria)蓝细菌门(Cyanobacteria)和拟杆菌门(Bacteroidetes),其中汇成河段水中的蓝细菌门(Cyanobacteria)、Chao1指数和Shannon指数最高,蓝细菌门(Cyanobacteria)从上游到下游显著增加,污水管网老化泄漏及点源排放是导致该河段Chao1指数和Shannon指数偏高的主要原因。冗余分析(RDA)表明总氮(TN)在三条城市内河河段的水生细菌群落分布中起关键作用,氨氮(NH3-N)和总磷(TP)以及叶绿素a(Chla)也在不同程度上影响着水体中细菌群落的组成。研究结果为精准高效保障城市内河水质提供了参考。Abstract: To explore planktonic bacteria community structure characteristics and driving factors of Wuhu city inland waters, three representative cities inland river (the central city in the river, the city river, south road section) were selected as the research objects. A total of 18 monitoring cross sections were setup according to the channel length and distribution of outlet. In this study, we analyzed the community structure characteristics of planktonic bacteria in three Wuhu inland rivers by16S rRNA high-throughput sequencing technologies, in September 2020. The results showed that Wuhu inland river had a highly abundant bacterial diversity. The main dominant bacteria species are similar, including Proteobacteria, Actinobacteria, Cyanobacteria and Bacteroidetes. Compared with the Zhongyangcheng reach and Zhongshan South Road reach, the number of Cyanobacteria in the confluence reach is significantly higher, and the number of Cyanobacteria in the confluence reach is significantly increased from upstream to downstream. The Chao1 index and Shannon index of water samples from the city river system are obviously higher than those from the Central City River system and the South Zhongshan Road River. This consequence may be resulted from peripheral sewage pipe network aging leakage and point source discharge. The result of redundancy analysis (RDA) showed that total nitrogen (TN) play key roles in the distribution of aquatic bacterial community in three urban inland rivers. And mmonia nitrogen (NH3-N), total phosphorus (TP) and chlorophyll a (Chla) also affect the composition of bacterial community. These findings may provide a reference for efficiently governing the water quality of city inland river.
-
Key words:
- Wuhu /
- urban inland river /
- community structure /
- biodiversity
-

-
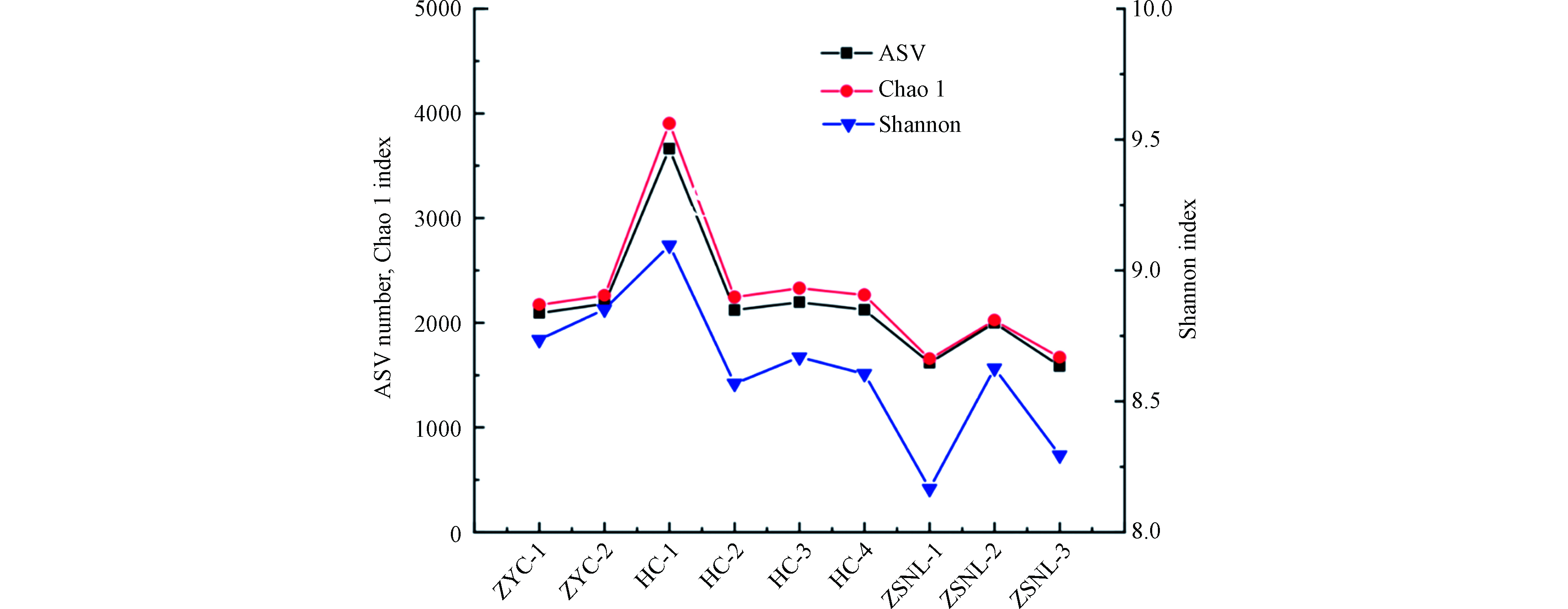
图 2 芜湖城市内河水体微生物多样性指数变化
Figure 2. Changes of microbial diversity index in Wuhu urban inland rivers

图 3 基于Bray-Curtis 距离的ASV水平的群落聚类分析
Figure 3. Principal coordinates analysis (PCoA) plot derived from the Bray-Curtis distance among seawater samples based on the occurrences and abundances of Amplicon Sequence Variants (ASVs )

图 4 芜湖城市内河门分类水平上细菌群落结构与分布图
Figure 4. Community structure and distribution of bacteria at the level of phylum classification in Wuhu inland rivers

图 5 芜湖城市内河属水平细菌群落结构与分布图
Figure 5. Community structure and distribution of bacteria at level of genus classification in Wuhu inland rivers

图 6 芜湖城市内河水体水样的优势菌门与环境因子的RDA3序图
Figure 6. RDA sequence diagram of dominant phyla and environmental factors in urban river water samples in Wuhu
表 1 采样断面经纬度坐标
Table 1. Longitude and latitude coordinates of river sections
河段
Reach采样点
Sampling sites微生物送检编号
Microbial inspection number经纬度
Longitude and latitude中央城河段 ZYC1 ZYC-1 E118°37′18.9″, N31°29′90.7″ ZYC2 ZYC-2 E118°37′22.1″, N31°29′63.4″ 汇成河段 HC1 HC1 E118°37′82.6″, N31°29′96.1″ HC2 HC-2 E118°37′82.2″, N31°29′94.3″ HC3 HC-3 E118°37′80.4″, N31°29′91.8″ HC5 HC-4 E118°37′77.2″, N31°29′64.4″ 中山南路河段 ZSNL2 ZSNL-1 E118°35′91.6″, N31°30′31.8″ ZSNL5 ZSNL-2 E118°35′89.6″, N31°30′23.9′ ZSNL8 ZSNL-3 E118°35′81.3″, N31°29′85.8″  下载: 导出CSV
下载: 导出CSV
表 2 芜湖城市内河水体理化性质
Table 2. physical and chemical properties of wuhu urban river water body
河段
Reach采样点
Sampling
sitespH EC* WT/
℃TP***/
(mg·L−1)TN*/
(mg·L−1)DO COD**/
(mg·L−1)BOD5 NH3-N*/
(mg·L−1)Chl a/
(μg·L−1)SD/
cmORP 中央城
河段ZYC1 8.05 423 28.0 0.170 4.07 2.7 8.85 3.80 0.58 30 48 162 ZYC2 7.81 796 27.4 0.153 3.48 1.6 12.70 8.89 1.70 15 54 174 ZYC3 7.79 434 25.0 0.221 4.41 2.4 4.71 3.30 2.20 16 91 167 汇成河段 HC1 7.81 435 28.2 0.179 5.16 2.3 12.10 8.47 1.54 82 49 153 HC2 7.85 460 28.9 0.163 5.69 4.6 13.80 9.66 2.18 44 46 143 HC3 7.76 406 28.7 0.176 5.56 5.1 10.90 7.63 2.44 33 41 204 HC4 7.97 466 28.8 0.173 5.93 5.8 19.90 13.93 3.12 30 43 171 HC5 8.88 516 29.0 0.236 4.42 9.1 18.40 2.90 3.34 31 43 138 HC6 8.49 449 28.9 0.258 5.31 9.1 16.20 2.80 3.44 40 54 146 HC7 7.75 438 28.4 0.273 5.08 5.4 8.60 6.02 3.54 55 57 183 中山南
路河段ZSNL1 7.57 647 31.3 0.305 7.50 2.6 13.30 9.31 2.34 31 113 194 ZSNL2 7.70 396 28.2 0.584 3.40 3.1 11.40 7.98 4.42 32 100 180 ZSNL3 7.73 407 28.4 0.387 5.48 4.2 15.10 10.50 4.14 23 121 171 ZSNL4 7.91 440 28.5 0.939 10.43 4.0 31.10 21.77 3.81 56 47 156 ZSNL5 9.05 420 30.0 0.927 9.33 7.7 31.50 3.50 8.64 157 45 128 ZSNL6 9.46 402 28.3 0.729 8.48 17.8 28.10 12.60 6.18 22 59 114 ZSNL7 9.43 424 30.5 0.762 8.14 17.1 36.40 11.50 6.30 115 55 137 ZSNL8 9.49 411 30.9 0.885 5.70 17.4 22.30 11.70 4.43 68 51 119 ZSNL9 9.24 420 30.4 0.708 6.38 15.1 24.50 8.80 3.88 42 70 127 注: * P<0.05,** P<0.01
下载: 导出CSV
表 3 城市黑臭水体分级标准
Table 3. Classification standards for urban black and odorous water bodies
特征指标
Characteristic index轻度黑臭
Mild black smelly重度黑臭
Heavy black smelly透明度/cm 25—10* <10* 溶解氧/(mg·L−1) 0.2—2.0 <0.2 氧化还原电位/mV −200—50 < −200 氨氮/(mg·L−1) 8.0—15 >15 注: * 水深不足 25 cm时,该指标按水深的 40%取值。
Note: * When the water depth is less than 25 cm, the value of this index is 40% of the water depth
下载: 导出CSV
表 4 研究区细菌群落的丰度和多样性
Table 4. Abundance and diversities of bacterial communities in the three inland rivers
样品
SampleASV数量*
ASVs门数量*
Phylum属数量**
GenusChao1指数*
Chao1 index观察到的物种*
Observed_Species香农指数*
Shannon index辛普森指数
Simpson indexZYC-1 2094 28 270 2172.36 2017.30 8.74 0.9907 ZYC-2 2181 20 226 2261.00 2140.40 8.85 0.9926 HC-1 3662 24 251 3901.71 3270.20 9.10 0.9894 HC-2 2122 22 259 2244.94 2050.80 8.57 0.9869 HC-3 2197 19 241 2330.21 2079.70 8.67 0.9873 HC-4 2125 17 216 2264.49 2061.00 8.60 0.9873 ZSNL-1 1618 20 188 1655.60 1573.60 8.17 0.9895 ZSNL-2 2000 20 262 2022.43 1992.20 8.63 0.9921 ZSNL-3 1587 22 239 1668.32 1515.40 8.29 0.9905 注: * P<0.05,** P<0.01
下载: 导出CSV
表 5 优势菌门与环境因子的 Pearson 相关性
Table 5. Pearson's correlation coefficient of predominant bacteriaat the phylum level and environmental factors
优势菌门
PhylumpH EC DO WT TP TN Chla NH3-N Proteobacteria 0.636 -0.288 −0.242 0.687* 0.886** 0.776* 0.802** 0.993** Actinobacteria −0.65 0.272 −0.099 −0.767* −0.624 −0.931** −0.811** 0.807** Cyanobacteria 0.069 0.01 0.413** 0.094 -0.514 0.031 −0.191 −0.401 Bacteroidetes −0.072 −0.17 −0.158 0.001 0.482 0.176 0.253 0.411 Verrucomicrobia −0.366 −0.115 0.791* −0.103 −0.495 −0.019 −0.101 −0.586 注: * P<0.05,** P<0.01
下载: 导出CSV
-
[1] 武福平, 夏传, 王艳琴, 等. 西北典型村镇集雨窖水水质变化及特性 [J]. 环境工程学报, 2014, 8(9): 3541-3545. WU F P, XIA C, WANG Y Q, et al. Quality and its variation of water in rainwater collection cellar of rural areas of northwest China [J]. Chinese Journal of Environmental Engineering, 2014, 8(9): 3541-3545(in Chinese).
[2] IBEKWE A M, MA J C, MURINDA S E. Bacterial community composition and structure in an Urban River impacted by different pollutant sources [J]. Science of the Total Environment, 2016, 566/567: 1176-1185. doi: 10.1016/j.scitotenv.2016.05.168 [3] SUTHAR S, SHARMA J, CHABUKDHARA M, et al. Water quality assessment of river Hindon at Ghaziabad, India: Impact of industrial and urban wastewater [J]. Environmental Monitoring and Assessment, 2010, 165(1/2/3/4): 103-112. [4] LUNDGAARD A, TREUSCH A H, STIEF P, et al. Nitrogen cycling and bacterial community structure of sinking and aging diatom aggregates [J]. Aquatic Microbial Ecology, 2017, 79(2): 85-99. doi: 10.3354/ame01821 [5] VINK R, BEHRENDT H, SALOMONS W. Development of the heavy metal pollution trends in several European rivers: An analysis of point and diffuse sources [J]. Water Science and Technology, 1999, 39(12): 215-223. doi: 10.2166/wst.1999.0549 [6] YUNKER M B, BACKUS S M, GRAF PANNATIER E, et al. Sources and significance of alkane and PAH hydrocarbons in Canadian arctic rivers [J]. Estuarine, Coastal and Shelf Science, 2002, 55(1): 1-31. doi: 10.1006/ecss.2001.0880 [7] GUO X P, LU D P, NIU Z S, et al. Bacterial community structure in response to environmental impacts in the intertidal sediments along the Yangtze Estuary, China [J]. Marine Pollution Bulletin, 2018, 126: 141-149. doi: 10.1016/j.marpolbul.2017.11.003 [8] 柴晓娟, 骆大伟, 吴春笃, 等. 水体中微生物分布及与环境因素的相关性研究 [J]. 人民长江, 2008, 39(3): 45-47,110. doi: 10.3969/j.issn.1001-4179.2008.03.017 CHAI X J, LUO D W, WU C D, et al. Research on relation of micro-organism distribution in water and environmental factor [J]. Yangtze River, 2008, 39(3): 45-47,110(in Chinese). doi: 10.3969/j.issn.1001-4179.2008.03.017
[9] SALA O E, CHAPIN F S, ARMESTO J J, et al. Global biodiversity scenarios for the year 2100 [J]. Science, 2000, 287(5459): 1770-1774. doi: 10.1126/science.287.5459.1770 [10] 陈兆进, 丁传雨, 朱静亚, 等. 丹江口水库枯水期浮游细菌群落组成及影响因素研究 [J]. 中国环境科学, 2017, 37(1): 336-344. doi: 10.3969/j.issn.1000-6923.2017.01.041 CHEN Z J, DING C Y, ZHU J Y, et al. Community structure and influencing factors of bacterioplankton during low water periods in Danjiangkou Reservoir [J]. China Environmental Science, 2017, 37(1): 336-344(in Chinese). doi: 10.3969/j.issn.1000-6923.2017.01.041
[11] LIU Z H, HUANG S B, SUN G P, et al. Phylogenetic diversity, composition and distribution of bacterioplankton community in the Dongjiang River, China [J]. FEMS Microbiology Ecology, 2012, 80(1): 30-44. doi: 10.1111/j.1574-6941.2011.01268.x [12] LOGUE J B, LANGENHEDER S, ANDERSSON A F, et al. Freshwater bacterioplankton richness in oligotrophic lakes depends on nutrient availability rather than on species–area relationships [J]. The ISME Journal, 2012, 6(6): 1127-1136. doi: 10.1038/ismej.2011.184 [13] 杨浩, 张国珍, 杨晓妮, 等. 16S rRNA高通量测序研究集雨窖水中微生物群落结构及多样性 [J]. 环境科学, 2017, 38(4): 1704-1716. YANG H, ZHANG G Z, YANG X N, et al. Microbial community structure and diversity in cellar water by 16S rRNA high-throughput sequencing [J]. Environmental Science, 2017, 38(4): 1704-1716(in Chinese).
[14] 谢建平, 韩玉波, 刘钢, 等. 2015年中国微生物遗传学研究领域若干重要进展 [J]. 遗传, 2016, 38(9): 765-790. XIE J P, HAN Y B, LIU G, et al. Research advances on microbial genetics in China in 2015 [J]. Hereditas, 2016, 38(9): 765-790(in Chinese).
[15] 刘永鑫, 秦媛, 郭晓璇, 等. 微生物组数据分析方法与应用 [J]. 遗传, 2019, 41(9): 845-862. LIU Y X, QIN Y, GUO X X, et al. Methods and applications for microbiome data analysis [J]. Hereditas, 2019, 41(9): 845-862(in Chinese).
[16] JI Y, ZHANGI J, HUANG X P, et al. Investigation and assessment of heavy metals in surface sediments of Ganjiang River, China [J]. Journal of Environmental Biology, 2014, 35(6): 1173-1179. [17] HAYDEN C J, BEMAN J M. Microbial diversity and community structure along a lake elevation gradient in Yosemite National Park, California, USA [J]. Environmental Microbiology, 2016, 18(6): 1782-1791. doi: 10.1111/1462-2920.12938 [18] 王元, 周俊芳, 韦信贤, 等. 海水和淡水养殖凡纳滨对虾肠道和鳃的菌群结构分析 [J]. 湖南农业大学学报(自然科学版), 2018, 44(2): 198-203. WANG Y, ZHOU J F, WEI X X, et al. Microbial community structure analysis of intestine and gill of Litopenaeus vannamei in seawater and freshwater [J]. Journal of Hunan Agricultural University (Natural Sciences), 2018, 44(2): 198-203(in Chinese).
[19] XIA N, XIA X H, LIU T, et al. Characteristics of bacterial community in the water and surface sediment of the Yellow River, China, the largest turbid river in the world [J]. Journal of Soils and Sediments, 2014, 14(11): 1894-1904. doi: 10.1007/s11368-014-0974-5 [20] YE L, ZHANG T. Bacterial communities in different sections of a municipal wastewater treatment plant revealed by 16S rDNA 454 pyrosequencing [J]. Applied Microbiology and Biotechnology, 2013, 97(6): 2681-2690. doi: 10.1007/s00253-012-4082-4 [21] 赵维, 王敬敬, 徐松, 等. 渤海湾表层海水中浮游细菌群落随离岸距离的分布特征及其影响因素 [J]. 海洋学报, 2019, 41(12): 156-171. ZHAO W, WANG J J, XU S, et al. Distribution characteristics and influencing factors of bacterioplankton community with offshore distance variation in the surface seawater of Bohai Bay [J]. Acta Oceanologica Sinica, 2019, 41(12): 156-171(in Chinese).
[22] MULKIDJANIAN A Y, KOONIN E V, MAKAROVA K S, et al. The cyanobacterial genome core and the origin of photosynthesis [J]. PNAS, 2006, 103(35): 13126-13131. doi: 10.1073/pnas.0605709103 [23] 程豹, 望雪, 徐雅倩, 等. 澜沧江流域浮游细菌群落结构特征及驱动因子分析 [J]. 环境科学, 2018, 39(8): 3649-3659. CHENG B, WANG X, XU Y Q, et al. Bacterioplankton community structure in the lancang river basin and the analysis of its driving environmental factors [J]. Environmental Science, 2018, 39(8): 3649-3659(in Chinese).
[24] 薛银刚, 刘菲, 江晓栋, 等. 太湖不同湖区冬季沉积物细菌群落多样性 [J]. 中国环境科学, 2018, 38(2): 719-728. doi: 10.3969/j.issn.1000-6923.2018.02.037 XUE Y G, LIU F, JIANG X D, et al. The diversity of bacterial communities in the sediment of different lake zones of Lake Taihu in winter [J]. China Environmental Science, 2018, 38(2): 719-728(in Chinese). doi: 10.3969/j.issn.1000-6923.2018.02.037
[25] 孙庆花, 于德爽, 张培玉, 等. 1株海洋异养硝化-好氧反硝化菌的分离鉴定及其脱氮特性 [J]. 环境科学, 2016, 37(2): 647-654. SUN Q H, YU D S, ZHANG P Y, et al. Identification and nitrogen removal characteristics of a heterotrophic nitrification-aerobic denitrification strain isolated from marine environment [J]. Environmental Science, 2016, 37(2): 647-654(in Chinese).
[26] ZHANG L, SHEN Z, FANG W K, et al. Composition of bacterial communities in municipal wastewater treatment plant [J]. Science of the Total Environment, 2019, 689: 1181-1191. doi: 10.1016/j.scitotenv.2019.06.432 [27] ZHANG L, ZHONG M M, LI X C, et al. River bacterial community structure and co-occurrence patterns under the influence of different domestic sewage types [J]. Journal of Environmental Management, 2020, 266: 110590. doi: 10.1016/j.jenvman.2020.110590 [28] XIE W, ZHANG C L, ZHOU X D, et al. Salinity-dominated change in community structure and ecological function of Archaea from the lower Pearl River to coastal South China Sea [J]. Applied Microbiology and Biotechnology, 2014, 98(18): 7971-7982. doi: 10.1007/s00253-014-5838-9 [29] ZHANG M L, YU N, CHEN L Q, et al. Structure and seasonal dynamics of bacterial communities in three urban rivers in China [J]. Aquatic Sciences, 2012, 74(1): 113-120. doi: 10.1007/s00027-011-0201-z [30] ZHANG Y, ZHAO Z H, DAI M H, et al. Drivers shaping the diversity and biogeography of total and active bacterial communities in the South China Sea [J]. Molecular Ecology, 2014, 23(9): 2260-2274. doi: 10.1111/mec.12739 [31] KORAJKIC A, PARFREY L W, MCMINN B R, et al. Changes in bacterial and eukaryotic communities during sewage decomposition in Mississippi river water [J]. Water Research, 2015, 69: 30-39. doi: 10.1016/j.watres.2014.11.003 [32] 王鹏, 陈波, 李传琼, 等. 赣江南昌段丰水期细菌群落特征 [J]. 中国环境科学, 2016, 36(8): 2453-2462. doi: 10.3969/j.issn.1000-6923.2016.08.027 WANG P, CHEN B, LI C Q, et al. Bacterial communities in Nanchang section of the Ganjiang River in wet seaon [J]. China Environmental Science, 2016, 36(8): 2453-2462(in Chinese). doi: 10.3969/j.issn.1000-6923.2016.08.027
[33] 刘鹏远, 陈庆彩, 胡晓珂. 渤海湾湾口表层沉积物中的核心细菌群落结构及其对环境因子的响应 [J]. 微生物学通报, 2018, 45(9): 1940-1955. LIU P Y, CHEN Q C, HU X K. Structure characteristics of core bacterial communities in surface sediments and analysis on their responses to environmental factors in the inlet of Bohai Bay [J]. Microbiology China, 2018, 45(9): 1940-1955(in Chinese).
[34] STALEY C, GOULD T J, WANG P, et al. Species sorting and seasonal dynamics primarily shape bacterial communities in the Upper Mississippi River [J]. Science of the Total Environment, 2015, 505: 435-445. doi: 10.1016/j.scitotenv.2014.10.012 [35] CAI X L, YAO L, SHENG Q Y, et al. Properties of bacterial communities attached to artificial substrates in a hypereutrophic urban river [J]. AMB Express, 2018, 8(1): 1-11. doi: 10.1186/s13568-017-0531-x [36] 赵海萍, 李清雪, 陶建华. 渤海湾浮游细菌分布特征及环境影响因素 [J]. 水资源保护, 2018, 34(5): 88-94. doi: 10.3880/j.issn.1004-6933.2018.05.14 ZHAO H P, LI Q X, TAO J H. Distribution characteristics of bacterioplankton in Bohai Bay and its environmental influence factors [J]. Water Resources Protection, 2018, 34(5): 88-94(in Chinese). doi: 10.3880/j.issn.1004-6933.2018.05.14
[37] YIN H Q, NIU J J, REN Y H, et al. An integrated insight into the response of sedimentary microbial communities to heavy metal contamination [J]. Scientific Reports, 2015, 5: 14266. doi: 10.1038/srep14266 [38] 杜宛璘, 孙金辉, 麦永湛, 等. 珠江下游浮游细菌群落结构的时空分布特征 [J]. 湖泊科学, 2020, 32(2): 380-394. doi: 10.18307/2020.0208 DU W L, SUN J H, MAI Y Z, et al. Spatial and temporal distribution characteristics of bacterioplankton community structure in the downstream of Pearl River [J]. Journal of Lake Sciences, 2020, 32(2): 380-394(in Chinese). doi: 10.18307/2020.0208
[39] WANG L, ZHANG J, LI H L, et al. Shift in the microbial community composition of surface water and sediment along an urban river [J]. Science of the Total Environment, 2018, 627: 600-612. doi: 10.1016/j.scitotenv.2018.01.203 [40] NIU L H, LI Y, WANG P F, et al. Altitude-scale variation in nitrogen-removal bacterial communities from municipal wastewater treatment plants distributed along a 3600 m altitudinal gradient in China[J]. Science of the Total Environment, 2016, 559: 38-44. -
 点击查看大图
点击查看大图

计量
- 文章访问数: 5126
- HTML全文浏览数: 5126
- PDF下载数: 120
- 施引文献: 0
