


 下载:
下载:
-
土壤环境对可持续农耕种植至关重要[1]. 因此,促进农田生态系统养分的良性循环并探究其驱动机制成为我国土壤肥力与养分循环研究的主要领域之一[2]. 沼液和生物质炭作为常见土壤改良剂在改善土壤健康,促进农业可持续发展并减少不利环境影响方面得到广泛关注[3]. 沼液作为沼气工程的副产物,富含多种植物所需的大量元素(氮、磷、钾)及微量元素(锰、铁、锌),并且元素形态主要以速效状态呈现,可以更好地被植物吸收[4]. 沼液全部或部分替代氮肥(以下简称沼液替代)作为缓解因化学氮肥施用导致的环境问题,提高耕层养分储量和供应能力的方式之一,已得到广泛认可[5]. 然而,沼液自身固液比低,农用过程中易导致土壤黏性降低,加速氮素淋失迁移及氨挥发和氮氧化物排放,在污染环境的同时造成沼液氮素利用率下降[6 − 7]. 生物质炭能够通过其表面羟基羧基等官能团来缓解氮素淋失,同时其自身不易被微生物分解,可以降低环境碳排放风险. 因此在沼液替代的同时添加生物质炭可以有效解决高频高量施沼所带来的环境问题[8].
土壤微生物被认为是调节碳、氮素循环和维持农田生态系统稳定性的主要驱动力[9]. 碳固定与氮代谢作为土壤系统中重要的2个能量代谢途径,可用于表达碳和氮的转化过程[10 − 12]. 系统阐释和评估农田土壤中碳固定、氮代谢相关微生物的丰度及群落组成,有助于判断有机物料施用对土壤碳、氮循环的影响. 已有研究结果表明,芽单胞菌属(Gemmatimonas)、绿弯菌属(Chloroflexi)、Gaiella和链霉菌属(Streptomyces)等微生物可作为土壤碳固定的标记性微生物种群,慢生根瘤菌(Bradyrhizobium)、硝化螺旋菌属(Nitrospira)和亚硝化螺菌属(Nitrosospira)等细菌可作为氮代谢的标记性微生物种群[13 − 20]. acsA、frdA和aclB等功能基因是碳固定过程的关键性基因,nirK3、nifH和nosZ2等基因对于研究氮代谢过程至关重要[21 − 25]. 有研究显示,土壤理化性质的改变会直接或间接地影响微生物的群落演替[26]. 不同类型微生物的丰富度决定微生物群落的功能,而功能基因丰度可以反映微生物群落功能对环境变化的响应[27]. 并且微生物对碳、氮元素的迁移,主要通过功能基因控制关键酶,实现调节碳、氮元素的循环过程. 因此,在预测微生物参与碳氮循环的潜力和代谢特征时,相关功能基因丰度成为关键因素[28]. Zhao等[29]研究表明,氮和碳循环基因的丰度增加,会影响土壤硝化潜力,微生物多样性与功能多样性增加,氮循环中微生物生物量、土壤养分含量和功能过程同时发生变化. 魏甲彬等[30]研究表明,添加生物质炭会增加根际土壤微生物群落的丰度,是评估土壤质量与植物生长方面的关键因素.
目前多数研究聚焦于单施沼液、生物质炭或与化肥联合施用对农田土壤养分及微生物组成的影响,而对生物质炭与沼液联合施用的研究较少,并且,缺乏长期施用生物质炭与沼液替代化肥对土壤微生态环境的动态影响. 据此本文以生物质炭与沼液替代化肥的旱地农田土壤为研究对象,揭示5年研究期内土壤养分含量随生物质炭与沼液替代氮肥的响应特征,并利用微生物宏基因组测序技术,研究生物质炭与沼液施入对碳固定、氮代谢途径微生物及功能基因的影响. 探究能否通过生物质炭与沼液替代化肥施用,实现可持续地提高旱地土壤性质与改善土壤微环境.
-
研究试验地点位于浙江省浙江科技大学(30°13′N,120°1′E). 近5年试验区年均降水量、年均日照时长和年均气温分别为
1573 mm、1970 h和17.8 ℃,气候类型属于亚热带季风气候. 供试土壤为典型南方黄红壤,基本理化值为:有机碳含量5.1g·kg−1,有机质含量8.79 g·kg−1,碱解氮含量45.87 mg·kg−1,氨态氮3.79 mg·kg−1,容重1.45 g·cm−3,全氮含量0.6 g·kg−1,pH 5.44,硝态氮含量3.54 mg·kg−1. -
供试的生物质炭制备原料来自浙江省衢州市顺康牧业有限公司,经浙江省金华市金锅公司以猪粪为原料,在350 ℃下厌氧裂解制备,理化性质为:有机碳含量605 g·kg−1,全氮含量3.95 g·kg−1,硝态氮11.37 mg·kg−1,pH 8.5,铵态氮含量15 mg·kg−1. 供试沼液原料取自顺康牧业有限公司的猪场废水,经发酵、沉淀、浓缩等处理,得到灌溉沼肥,其中重金属含量分别为:总锌(Zn)≤ 1.33 mg·L−1、总铅(Pb)≤ 0.01 mg·L−1、总铬(Cr)≤ 0.127 mg·L−1均符合国家标准(GB/T
40750 —2021). 沼液理化性质为:硝态氮含量10 mg·L−1,pH 8.5,铵态氮含量220 mg·L−1,有机质含量4.04 g·L−1,全氮含量2.38 g·L−1. -
实验于2018年12月—2023年4月间进行,实验种植农作物为包菜,包菜于前一年12月15日播种,次年4月30日收获. 5年间施肥方式保持一致,各小区面积12 m2(3 m×4 m). 在实验设计前,对当地包菜的常规施肥量进行了调查,得出化肥N施用量为225 kg·hm−2. 基于氮磷钾总用量不变,按每小区氮含量折算沼液替代氮肥用量,尿素(N,46%):氯化钾:钙镁磷肥养分质量比为 1:0.54:1.04. 其中替代处理的磷钾含量不足部分通过添加氯化钾和钙镁磷肥补齐. 实验共设计5个处理,其中,常规施肥(C0B0)、100%(等养分)沼液替代氮肥(C0B1)、2%(土壤质量比)生物质炭(C1B0)和100%沼液替代氮肥+2%生物质炭(C1B1),同时设置不施肥处理(CK)为对照,每个处理3个重复,共计15个小区. 包菜种植规格保持各小区植株行距(40 cm)与株距(20 cm)均匀分布. 播种前两周去除地表杂草及凋谢物,通过人工散施,将生物质炭一次性均匀施入各自对应小区(45 t·hm−2),再采用人工翻耕的方式,翻耕至土层深度为10—20 cm,此后不再另施生物质炭. 后期播种前都需要再次施加沼液和肥料,且每季保持相同施用量(79.6 m3·hm−2),沼液为每隔1个月施用1次,共施用3次.
-
测试土壤于每年包菜收获前1周采集1次,5年间共采集5次,每次按照S形土壤采集法,采集土层0—20 cm的土壤样本,并放于4 ℃恒温箱中带回实验室. 及时剔除植物根系及凋落物等杂质,将样品均匀分成两份. 一份自然风干,研钵研磨后过筛,用于测定土壤基本理化性质,测试方法参考《土壤农业化学分析方法》[31]. 一份储存于-80 ℃冰箱中,用于测定土壤微生物丰度与功能基因.
-
利用pH计电位法测定各小区土壤pH(水土比5:1). 全碳、全氮利用元素分析仪(Elementar Vario EL cube)测定. 有机碳与有机质采用重铬酸钾外加热法测定. 硝态氮与铵态氮采用浸提法测定. 碱解氮采用碱解扩散法测定.
-
使用试剂盒(FastLee DNA kit,中国杭州力盟生物医药科技有限公司)提取土壤DNA ,通过PCR扩增细菌16S核糖体RNA基因的V3—V4区域,加入试剂进行PCR反应,再从2%琼脂糖凝胶中提取扩增子,并使用AxyPrep DNA凝胶提取试剂盒(Axygen Biosciences,Union City,CA,U.S.)提纯,使用QuantiFluor™-ST(Promega,U.S.)定量. 通过Qubit®3.0(Life Invitrogen)定量纯化的PCR产物,并将条形码不同的每24个扩增子均等混合. 合并的DNA产物用于按照Illumina的基因组DNA文库制备程序构建Illumina Pair-End文库. 使用USEARCH将相似性≥97%的序列进行OTU(operational taxonomic units)聚类,通过使用UCLUST v1.2.22q对Silva 16S rRNA数据库对比,进行OTU物种注释,置信度(阈值)为70%,并对不同物种分类水平下统计每个样本的群落组成.
-
使用土壤DNA快速提取试剂盒(TIANNAMP Soil DNA Kit,中国Tian-Gen公司)提取土壤中微生物总DNA,利用NanoDrop-2000(Thermo Fisher Scientific,MA,USA)检测提取的DNA纯度,DNA浓度的检测使用TBS-380检测. 采用高通量自动微量加样设备分别将样品板和引物板试剂添加至高通量 qPCR 芯片的纳米孔中,在SmartChip Real-Time PCR System中实行qPCR反应及荧光信号检测,并自动生成扩增曲线和溶解曲线. 通过样本的检出情况和Ct值进行质控、标准化得到各基因在各样本中相对定量信息,Roche仪器检测获得16S rRNA基因的绝对定量,经公式:16S rRNA相对定量/16 rRNA绝对定量=基因相对定量/基因绝对定量,换算获得其他基因的绝对定量信息[32 − 33]. 从基因目录出发,使用blastp(Version 2.3.0)将基因集序列与NR数据库进行比对,获得碳固定微生物菌属(Gemmatimonas、Chloroflexi、Gaiella和Streptomyces等)和氮代谢微生物菌属(Bradyrhizobium、Nitrospira和Nitrosospira等)的注释信息,并结合基因丰度,获得不同分类层级微生物物种丰度.
-
实验数据采集从2019年4月至2023年4月,将5年的采集数据求平均值并进行分析. 统计与分类采用Excel 2013. 利用SPSS 27进行单因素(one-way ANOVA)和Duncan法进行方差分析和多重比较(α=0.05),差异性显著采用P<0.05水平标注. 用Pearson法对微生物和功能基因进行相关分析. 使用 Origin 2021进行绘图与添加显著性标签.
-
从图1可以看出,生物质炭与沼液施用对旱地表层土壤理化性质具有显著影响. 与CK和C0B0处理相比,生物质炭与沼液无论是单施还是联合施用均能显著提高土壤pH,其中C0B1、C1B0与C1B1分别提高7.3%、33.1%和35.9%,这可能是因所施用的生物质炭、沼液均采用动物源有机废物制备而成,整体呈碱性,通过酸碱调节,降低了土壤酸化的风险,这一结果与覃潇敏等[34]研究一致. 对全量养分分析,添加生物质炭的处理显著高于未添加生物质炭的处理,其中C1B1处理中的总碳与总氮含量增幅最为显著,分别为CK的3.4倍与3.5倍,这可能是由于外部碳源与氮源的引入导致的碳氮养分含量的增加[35]. 相比于C0B0处理,C0B1处理中有机碳和硝态氮含量显著增加了16.8%和77.5%,其主要原因可能是沼液中富含多种氮素养分,通过植物根系吸收和土壤固持,导致氮素养分的增加,这一结果与温云杰等[36]利用沼液替代化肥施用对大葱(Allium.fistulosum L.)种植研究保持一致. 而C1B0处理中铵态氮含量相比于C0B0与C0B1处理,均有显著增加,分别增加了30.7%与21.1%. 碳沼联合施用(C1B1)比单施生物质炭(C1B0)处理,对有机碳、铵态氮与硝态氮养分含量提升效果更为显著,分别增加了39.0%、68.4%与42.7%,这可能是由于生物质炭对固液比低的沼液进行了吸附,减少了沼液的挥发损失,导致土壤中碳氮养分的增加[37]. 因此,长期生物质炭与沼液的连续施用对土壤养分含量以及土质性能有显著改善作用.
-
通过对土壤微生物群落丰度的检测,得到不同处理下土壤细菌丰度情况,将相对丰度超过1%的微生物定义为优势菌. 从图2a可以看出,5个处理的优势菌门相同,主要为:变形菌门(Proteobacteria)、绿弯菌门(Chloroflexi)、放线菌门(Actinobacteria)、酸杆菌门(Acidobacteria)、浮霉菌门(Planctomycetes)、芽单胞菌门(Gemmatimonadetes)、蓝光合菌门(Cyanobacteria)和装甲菌门(Armatimonadetes)共8个,占总菌门91.8%—97.7%. 相比于CK处理,C0B0处理显著增加Cyanobacteria的相对丰度,而显著降低Proteobacteria、Gemmatimonadetes和Armatimonadetes的相对丰度,C0B1、C1B0和C1B1处理显著增加Chloroflexi、Actinobacteria和Acidobacteria的相对丰度,显著降低Cyanobacteria与Armatimonadetes的相对丰度.
由图2b可知,各处理中碳固定优势菌属为Gemmatimonas、未分类的厌氧绳菌属(Anaerolineaceae_uncultured)、未分类的绿弯菌属(Roseiflexaceae_uncultured)、假诺卡氏菌属(Pseudonocardia)和Streptomyces. 且不同处理间碳固定优势菌属丰度差异显著,其中碳固定优势菌属丰度在C1B1中达最高,相比于CK处理,增涨64%. 与CK处理相比较,单独施加化肥、生物质炭和炭-沼液联合施用(C1B1)的处理中Gemmatimonas丰度显著降低,分别降低了39.0%、55.4%和51.3%. 生物质炭的单施或与沼液混施对Anaerolineaceae_uncultured和Roseiflexaceae_uncultured菌属的相对丰度有显著提高,单施生物质炭时提高168.7%和156.1%,生物质炭和沼液联合施用时提高246.4%和220.2%. Pseudonocardia菌属在单施沼液处理中显著降低18.2%,施加生物质炭的处理中则显著增加,其中C1B0与C1B1分别增加43.7%和50.3%. C1B1中Streptomyces菌属丰度最高,相比于CK与C0B0处理,分别提高74.0%和65.6%,这可能有两方面原因;一方面是生物质炭通过吸附沼液中的养分,储存于生物质炭的孔隙中,缓解沼液在高温条件下的挥发损失,提供了充足的养料,促进土壤中的菌群丰度的增加. 另一方面可能是生物质炭的施用导致土壤pH值升高,使得菌群的生存环境得到改善,再联合沼液施用,增加土壤养分含量,提高土壤支持多种生态系统功能的能力,最终导致菌群丰度的显著提升[38].
由图2c可知,分枝杆菌属(Mycobacterium)、Bradyrhizobium、Nitrospira、小梨形菌属(Pirellula)和根瘤菌(Rhizobium)为氮代谢优势菌属,且优势菌属丰度在不同处理间差异达显著水平,且生物质炭与沼液联合施用处理中丰度达到最高值,相比于CK处理,增涨65.7%. 施用化肥、沼液或生物质炭的处理中Mycobacterium菌属丰度相比于CK处理均有显著提升,其中沼液单独施用提升最显著,达到139.4%. 此外,单施沼液处理中Rhizobium菌属丰度比单施化肥处理增加了20.8%,这可能是由于生物质炭的施用刺激了固氮菌的活性,并且通过利用沼液中丰富的养分实现丰度的增加[39 − 40]. Bradyrhizobium菌属丰度在生物质炭单施或与沼液配施时显著增加35.1%和41.8%,而单施化肥或沼液的处理中显著降低了57.2%和51.8%. 生物质炭和沼液配施的处理中Nitrospira和Pirellula菌群丰度显著高于其他处理,相比于CK处理显著提升148.7%和115.7%,这一方面可能是由于生物质炭的多孔隙结构增加了土壤的透气性,提高了土壤中氧气含量,引起好氧菌丰度的增加[41]. 另一方面可能是由于沼液施用引入了丰富的氮素养分,为硝化作用提供了充足的原料[42]. 由此说明,长期使用生物质炭与沼液,有利于维护土壤微生态环境的健康稳定,对土壤碳氮循环产生有利影响.
-
利用荧光定量RCR法对土壤功能基因测定,注释到13种碳固定基因和22种氮代谢基因. 对比基因测试结果,选取基因丰度排名前10的功能基因进行组间分析. 通过碳固定基因结果可知(图3a),相较于CK与C0B0处理,C1B1的中的基因相对丰度显著增加,其中acsE相对丰度分别提高102.2%与97.4%,rbcL基因分别提高175.8%和95.3%. 沼液或生物质炭的单独施用,对accA、acsA、acsE、rbcL、cdaR、korA、frdA、pccA、acsB的基因相对丰度显著高于单施化肥,其中C0B1和C1B0中acsE的丰度分别提高35.4%与47.8%,这一方面可能是由于生物质炭自身的难降解性导致土壤碳库的增加,间接有利于土壤的固碳[43]. 另一方面,沼液施用提高了土壤中养分浓度,改变了微生物的代谢途径[44]. 对氮代谢基因检测结果进行组间分析(图3b),其中硝化型功能基因为amoB、amoA2、nxrA,反硝化型功能基因为narG、napA、nirK3、nosZ2、nirS1,氮矿化型功能基因gdhA,固氮型功能基因nifH. 与CK相比较,C0B0、C0B1、C1B0和C1B1处理nxrA、narG、napA、nirK3与gdhA功能基因丰度显著提高,这可能是由于生物质炭与沼液施用,引入了大量的活性微生物,创建了一个富营养化的微环境领域,补充了硝化细菌、反硝化细菌和矿化细菌所需的底物,刺激了氮代谢细菌的繁殖,导致基因丰度的增加[45]. 与C0B0处理相比较,C0B1处理nifH、nxrA、nirK3基因相对丰度显著增加,C1B0处理中nxrA、narG、napA、nirS1基因丰度有显著增加,C1B1处理中硝化型基因nxrA显著增加54.0%,反硝化基因narG、napA、nirK3、nirS1也显著增加,这与郑一玲等[46]研究一致. 这可能是由于外部碳源与氮源的加入,为土壤微生物提供富足的养分环境,使得微生物通过调整基因表达来适应环境变化,进而提高功能基因的表达[47]. 总体分析可知,长期生物质炭与沼液替代化肥施用,可促进土壤碳固定与氮代谢途径基因的表达,提升土壤微生物参与碳氮循环潜力和代谢特征.
-
为了解碳固定与氮代谢微生物的生物学特性与生态学适应性,选取丰度排名前十的微生物(属水平)及关键型功能基因进行相关性分析. 由图4a可知,碳固定功能基因与微生物之间正相关性(85%)的占比大于负相关性(15%),10种菌属中Vicinamibacterales_norank、Streptomyces和Gaiella与功能基因如accA、acsE、frdA、cdaR和pccA呈显著正相关,表明3种细菌在碳固定途径中起到重要的固碳作用. 此外rbcL基因与Vicinamibacterales_norank和Streptomyces菌属也存在显著正相关. 并且frdA和pccA基因与菌属Anaerolineaceae_uncultured、Roseiflexaceae_uncultured、Pseudonocardia和Nocardioides呈显著正相关. 基因pccA与RB41呈显著正相关.
由图4b可知,氮代谢关键基因与主要微生物之间正相关性(73%)的占比大于负相关性(27%). 其中硝化型基因(nxrA)和反硝型基因(nirK3)与Frankiales_norank、Nitrosospira、Amycolatopsis和Pirellula菌属之间显著正相关. 反硝化基因(napA和nirS1)与Nitrospira、Pirellula和Bacillus之间显著正相关. Nitrosomonas、Nitrospira、Amycolatopsis和Pirellula4种菌属与反硝化型基因(narG)显著正相关. Rhizobium与氮固定型基因(nifH)显著正相关. Bacillus菌属与nirK3显著正相关,而与有氧氨氧化型基因(amoA2)显著负相关. 此外,amoA2还与Pirellula和Nitrospira显著负相关. 反硝化型基因(nosZ2)与Pirellula、Amycolatopsis和Nitrosomonas3种菌属显著负相关.
-
采取生物质炭与沼液联合施用可显著增加土壤有机碳、全碳、全氮、铵态氮、硝态氮含量并提高土壤pH. 有益于碳固定与氮代谢微生物属水平种群丰度的增加,可以提高固碳功能基因与氮代谢功能基因表达水平,增强土壤微生物的固碳与反硝化能力,促进土壤系统的碳、氮循环. 因此采取生物质炭与沼液联合替代化肥施用,有利于维护土壤微环境健康的同时增强微生物的生态适应性,保障土地生产力的稳定.
生物质炭-沼液联用对土壤微生物及碳氮功能基因的影响
Effects of biochar-biogas slurry application on soil microorganisms and functional genes
-
摘要: 土壤微生物是土壤养分的重要驱动力,明确生物质炭-沼液配施条件下土壤微环境响应特征对改善土壤健康、提升土壤质量和增强土壤可持续耕作能力具有重要意义. 本研究以动物源有机物料制成生物质炭和沼液为材料,连续种植5年的旱地黄红壤为研究对象,设计常规施肥(C0B0)、沼液替代氮肥(C0B1)、2%生物质炭(C1B0)和100%沼液替代氮肥+2%生物质炭(C1B1),同时设置不施肥处理(CK)为对照,以探究生物质炭与沼液替代化肥施用对旱地土壤肥力、碳固定和氮代谢途径微生物群落及功能基因的影响. 结果表明,相比于CK和C0B0处理,生物质炭与沼液无论是单施还是联合施用均能显著提高土壤pH,其中C1B1提升最为显著,增幅达35.9%. C1B1处理中的总碳与总氮含量提升效果最为显著,分别为CK的3.4倍与3.5倍. 并且C1B1处理中有机碳含量比C1B0处理显著增加39%. C0B1处理中硝态氮含量比C0B0处理显著增加77.5%. C1B0处理中铵态氮的含量比C0B1处理显著增加21.1%. 碳固定与氮代谢属水平优势菌属丰度在C1B1条件下最高,相比于CK处理,分别增加64.0%与65.7%. 与C0B0相比,C1B1显著增加碳固定和氮代谢基因丰度,其中acsE与nxrA相对丰度增加97.4%与54.0%. 碳固定菌属与功能基因pccA和frdA显著正相关,氮代谢菌属与nxrA、narG、napA、nirK3和nirS1功能基因显著正相关,而与amoA2和nosZ2显著负相关. 综上,生物质炭与沼液联合替代化肥施用可显著提升土壤养分含量,促进土壤微生物量增加,提高碳固定与氮代谢相关基因丰度,有益于调节土壤微系统健康.Abstract: Soil microorganisms are key drivers of soil nutrient cycling. Understanding the response characteristics of the soil microenvironment under the combined application of biochar and biogas slurry is crucial for improving soil health, enhancing soil quality, and increasing the sustainability of soil for agricultural use. This study used biochar made from animal-derived organic materials and biogas slurry, with a focus on a dryland red-yellow soil cultivated for five consecutive years. The experimental design included conventional fertilization (C0B0), biogas slurry substituting for nitrogen fertilizer (C0B1), 2% biochar (C1B0), and a combined application of 100% biogas slurry substituting for nitrogen fertilizer plus 2% biochar (C1B1), alongside a control with no fertilization (CK). The aim was to explore the effects of biochar and biogas slurry, as substitutes for chemical fertilizers, on soil fertility, carbon sequestration, and the microbial communities and functional genes involved in nitrogen metabolism in dryland soils. The results indicated that compared to the CK and C0B0 treatments, both the separate and combined applications of biochar and biogas slurry significantly increased soil pH, with the C1B1 treatment showing the most substantial increase, up to 35.9%. The C1B1 treatment also demonstrated the most notable increase in total carbon and total nitrogen contents, being 3.4 times and 3.5 times higher than those of CK, respectively. Additionally, the organic carbon content in C1B1 was significantly higher by 39% compared to C1B0. In the C0B1 treatment, nitrate nitrogen content increased significantly by 77.5% compared to C0B0, while ammonium nitrogen content in the C1B0 treatment increased significantly by 21.1% compared to C0B1. The highest abundance of dominant microbial genera associated with carbon fixation and nitrogen metabolism was observed under C1B1 conditions, with increases of 64.0% and 65.7%, respectively, compared to CK. Relative to C0B0, C1B1 significantly increased the abundance of genes associated with carbon fixation and nitrogen metabolism, with acsE and nxrA relative abundances increasing by 97.4% and 54.0%, respectively. The abundance of carbon-fixing genera was significantly positively correlated with the functional genes pccA and frdA, while nitrogen-metabolizing genera showed a significant positive correlation with nxrA, narG, napA, nirK3, and nirS1, and a significant negative correlation with amoA2 and nosZ2. In summary, the combined application of biochar and biogas slurry as a substitute for chemical fertilizers can significantly enhance soil nutrient content, promote microbial biomass, and increase the abundance of genes related to carbon fixation and nitrogen metabolism, thereby contributing to the regulation of soil microecosystem health.
-
Key words:
- biochar /
- biogas slurry /
- carbon fixation /
- nitrogen metabolism /
- microbial communities /
- functional genes.
-
土壤环境对可持续农耕种植至关重要[1]. 因此,促进农田生态系统养分的良性循环并探究其驱动机制成为我国土壤肥力与养分循环研究的主要领域之一[2]. 沼液和生物质炭作为常见土壤改良剂在改善土壤健康,促进农业可持续发展并减少不利环境影响方面得到广泛关注[3]. 沼液作为沼气工程的副产物,富含多种植物所需的大量元素(氮、磷、钾)及微量元素(锰、铁、锌),并且元素形态主要以速效状态呈现,可以更好地被植物吸收[4]. 沼液全部或部分替代氮肥(以下简称沼液替代)作为缓解因化学氮肥施用导致的环境问题,提高耕层养分储量和供应能力的方式之一,已得到广泛认可[5]. 然而,沼液自身固液比低,农用过程中易导致土壤黏性降低,加速氮素淋失迁移及氨挥发和氮氧化物排放,在污染环境的同时造成沼液氮素利用率下降[6 − 7]. 生物质炭能够通过其表面羟基羧基等官能团来缓解氮素淋失,同时其自身不易被微生物分解,可以降低环境碳排放风险. 因此在沼液替代的同时添加生物质炭可以有效解决高频高量施沼所带来的环境问题[8].
土壤微生物被认为是调节碳、氮素循环和维持农田生态系统稳定性的主要驱动力[9]. 碳固定与氮代谢作为土壤系统中重要的2个能量代谢途径,可用于表达碳和氮的转化过程[10 − 12]. 系统阐释和评估农田土壤中碳固定、氮代谢相关微生物的丰度及群落组成,有助于判断有机物料施用对土壤碳、氮循环的影响. 已有研究结果表明,芽单胞菌属(Gemmatimonas)、绿弯菌属(Chloroflexi)、Gaiella和链霉菌属(Streptomyces)等微生物可作为土壤碳固定的标记性微生物种群,慢生根瘤菌(Bradyrhizobium)、硝化螺旋菌属(Nitrospira)和亚硝化螺菌属(Nitrosospira)等细菌可作为氮代谢的标记性微生物种群[13 − 20]. acsA、frdA和aclB等功能基因是碳固定过程的关键性基因,nirK3、nifH和nosZ2等基因对于研究氮代谢过程至关重要[21 − 25]. 有研究显示,土壤理化性质的改变会直接或间接地影响微生物的群落演替[26]. 不同类型微生物的丰富度决定微生物群落的功能,而功能基因丰度可以反映微生物群落功能对环境变化的响应[27]. 并且微生物对碳、氮元素的迁移,主要通过功能基因控制关键酶,实现调节碳、氮元素的循环过程. 因此,在预测微生物参与碳氮循环的潜力和代谢特征时,相关功能基因丰度成为关键因素[28]. Zhao等[29]研究表明,氮和碳循环基因的丰度增加,会影响土壤硝化潜力,微生物多样性与功能多样性增加,氮循环中微生物生物量、土壤养分含量和功能过程同时发生变化. 魏甲彬等[30]研究表明,添加生物质炭会增加根际土壤微生物群落的丰度,是评估土壤质量与植物生长方面的关键因素.
目前多数研究聚焦于单施沼液、生物质炭或与化肥联合施用对农田土壤养分及微生物组成的影响,而对生物质炭与沼液联合施用的研究较少,并且,缺乏长期施用生物质炭与沼液替代化肥对土壤微生态环境的动态影响. 据此本文以生物质炭与沼液替代化肥的旱地农田土壤为研究对象,揭示5年研究期内土壤养分含量随生物质炭与沼液替代氮肥的响应特征,并利用微生物宏基因组测序技术,研究生物质炭与沼液施入对碳固定、氮代谢途径微生物及功能基因的影响. 探究能否通过生物质炭与沼液替代化肥施用,实现可持续地提高旱地土壤性质与改善土壤微环境.
1. 材料和方法(Materials and methods)
1.1 实验区概况
研究试验地点位于浙江省浙江科技大学(30°13′N,120°1′E). 近5年试验区年均降水量、年均日照时长和年均气温分别为
1573 mm、1970 h和17.8 ℃,气候类型属于亚热带季风气候. 供试土壤为典型南方黄红壤,基本理化值为:有机碳含量5.1g·kg−1,有机质含量8.79 g·kg−1,碱解氮含量45.87 mg·kg−1,氨态氮3.79 mg·kg−1,容重1.45 g·cm−3,全氮含量0.6 g·kg−1,pH 5.44,硝态氮含量3.54 mg·kg−1.1.2 实验材料
供试的生物质炭制备原料来自浙江省衢州市顺康牧业有限公司,经浙江省金华市金锅公司以猪粪为原料,在350 ℃下厌氧裂解制备,理化性质为:有机碳含量605 g·kg−1,全氮含量3.95 g·kg−1,硝态氮11.37 mg·kg−1,pH 8.5,铵态氮含量15 mg·kg−1. 供试沼液原料取自顺康牧业有限公司的猪场废水,经发酵、沉淀、浓缩等处理,得到灌溉沼肥,其中重金属含量分别为:总锌(Zn)≤ 1.33 mg·L−1、总铅(Pb)≤ 0.01 mg·L−1、总铬(Cr)≤ 0.127 mg·L−1均符合国家标准(GB/T
40750 —2021). 沼液理化性质为:硝态氮含量10 mg·L−1,pH 8.5,铵态氮含量220 mg·L−1,有机质含量4.04 g·L−1,全氮含量2.38 g·L−1.1.3 实验设计
实验于2018年12月—2023年4月间进行,实验种植农作物为包菜,包菜于前一年12月15日播种,次年4月30日收获. 5年间施肥方式保持一致,各小区面积12 m2(3 m×4 m). 在实验设计前,对当地包菜的常规施肥量进行了调查,得出化肥N施用量为225 kg·hm−2. 基于氮磷钾总用量不变,按每小区氮含量折算沼液替代氮肥用量,尿素(N,46%):氯化钾:钙镁磷肥养分质量比为 1:0.54:1.04. 其中替代处理的磷钾含量不足部分通过添加氯化钾和钙镁磷肥补齐. 实验共设计5个处理,其中,常规施肥(C0B0)、100%(等养分)沼液替代氮肥(C0B1)、2%(土壤质量比)生物质炭(C1B0)和100%沼液替代氮肥+2%生物质炭(C1B1),同时设置不施肥处理(CK)为对照,每个处理3个重复,共计15个小区. 包菜种植规格保持各小区植株行距(40 cm)与株距(20 cm)均匀分布. 播种前两周去除地表杂草及凋谢物,通过人工散施,将生物质炭一次性均匀施入各自对应小区(45 t·hm−2),再采用人工翻耕的方式,翻耕至土层深度为10—20 cm,此后不再另施生物质炭. 后期播种前都需要再次施加沼液和肥料,且每季保持相同施用量(79.6 m3·hm−2),沼液为每隔1个月施用1次,共施用3次.
1.4 土壤样品采集
测试土壤于每年包菜收获前1周采集1次,5年间共采集5次,每次按照S形土壤采集法,采集土层0—20 cm的土壤样本,并放于4 ℃恒温箱中带回实验室. 及时剔除植物根系及凋落物等杂质,将样品均匀分成两份. 一份自然风干,研钵研磨后过筛,用于测定土壤基本理化性质,测试方法参考《土壤农业化学分析方法》[31]. 一份储存于-80 ℃冰箱中,用于测定土壤微生物丰度与功能基因.
1.5 土壤理化性质的测定
利用pH计电位法测定各小区土壤pH(水土比5:1). 全碳、全氮利用元素分析仪(Elementar Vario EL cube)测定. 有机碳与有机质采用重铬酸钾外加热法测定. 硝态氮与铵态氮采用浸提法测定. 碱解氮采用碱解扩散法测定.
1.6 土壤微生物丰度的测定
使用试剂盒(FastLee DNA kit,中国杭州力盟生物医药科技有限公司)提取土壤DNA ,通过PCR扩增细菌16S核糖体RNA基因的V3—V4区域,加入试剂进行PCR反应,再从2%琼脂糖凝胶中提取扩增子,并使用AxyPrep DNA凝胶提取试剂盒(Axygen Biosciences,Union City,CA,U.S.)提纯,使用QuantiFluor™-ST(Promega,U.S.)定量. 通过Qubit®3.0(Life Invitrogen)定量纯化的PCR产物,并将条形码不同的每24个扩增子均等混合. 合并的DNA产物用于按照Illumina的基因组DNA文库制备程序构建Illumina Pair-End文库. 使用USEARCH将相似性≥97%的序列进行OTU(operational taxonomic units)聚类,通过使用UCLUST v1.2.22q对Silva 16S rRNA数据库对比,进行OTU物种注释,置信度(阈值)为70%,并对不同物种分类水平下统计每个样本的群落组成.
1.7 土壤碳氮循环功能基因测定
使用土壤DNA快速提取试剂盒(TIANNAMP Soil DNA Kit,中国Tian-Gen公司)提取土壤中微生物总DNA,利用NanoDrop-2000(Thermo Fisher Scientific,MA,USA)检测提取的DNA纯度,DNA浓度的检测使用TBS-380检测. 采用高通量自动微量加样设备分别将样品板和引物板试剂添加至高通量 qPCR 芯片的纳米孔中,在SmartChip Real-Time PCR System中实行qPCR反应及荧光信号检测,并自动生成扩增曲线和溶解曲线. 通过样本的检出情况和Ct值进行质控、标准化得到各基因在各样本中相对定量信息,Roche仪器检测获得16S rRNA基因的绝对定量,经公式:16S rRNA相对定量/16 rRNA绝对定量=基因相对定量/基因绝对定量,换算获得其他基因的绝对定量信息[32 − 33]. 从基因目录出发,使用blastp(Version 2.3.0)将基因集序列与NR数据库进行比对,获得碳固定微生物菌属(Gemmatimonas、Chloroflexi、Gaiella和Streptomyces等)和氮代谢微生物菌属(Bradyrhizobium、Nitrospira和Nitrosospira等)的注释信息,并结合基因丰度,获得不同分类层级微生物物种丰度.
1.8 数据处理及分析
实验数据采集从2019年4月至2023年4月,将5年的采集数据求平均值并进行分析. 统计与分类采用Excel 2013. 利用SPSS 27进行单因素(one-way ANOVA)和Duncan法进行方差分析和多重比较(α=0.05),差异性显著采用P<0.05水平标注. 用Pearson法对微生物和功能基因进行相关分析. 使用 Origin 2021进行绘图与添加显著性标签.
2. 结果与讨论(Results and discussion)
2.1 生物质炭与沼液施用对土壤理化性质的影响
从图1可以看出,生物质炭与沼液施用对旱地表层土壤理化性质具有显著影响. 与CK和C0B0处理相比,生物质炭与沼液无论是单施还是联合施用均能显著提高土壤pH,其中C0B1、C1B0与C1B1分别提高7.3%、33.1%和35.9%,这可能是因所施用的生物质炭、沼液均采用动物源有机废物制备而成,整体呈碱性,通过酸碱调节,降低了土壤酸化的风险,这一结果与覃潇敏等[34]研究一致. 对全量养分分析,添加生物质炭的处理显著高于未添加生物质炭的处理,其中C1B1处理中的总碳与总氮含量增幅最为显著,分别为CK的3.4倍与3.5倍,这可能是由于外部碳源与氮源的引入导致的碳氮养分含量的增加[35]. 相比于C0B0处理,C0B1处理中有机碳和硝态氮含量显著增加了16.8%和77.5%,其主要原因可能是沼液中富含多种氮素养分,通过植物根系吸收和土壤固持,导致氮素养分的增加,这一结果与温云杰等[36]利用沼液替代化肥施用对大葱(Allium.fistulosum L.)种植研究保持一致. 而C1B0处理中铵态氮含量相比于C0B0与C0B1处理,均有显著增加,分别增加了30.7%与21.1%. 碳沼联合施用(C1B1)比单施生物质炭(C1B0)处理,对有机碳、铵态氮与硝态氮养分含量提升效果更为显著,分别增加了39.0%、68.4%与42.7%,这可能是由于生物质炭对固液比低的沼液进行了吸附,减少了沼液的挥发损失,导致土壤中碳氮养分的增加[37]. 因此,长期生物质炭与沼液的连续施用对土壤养分含量以及土质性能有显著改善作用.
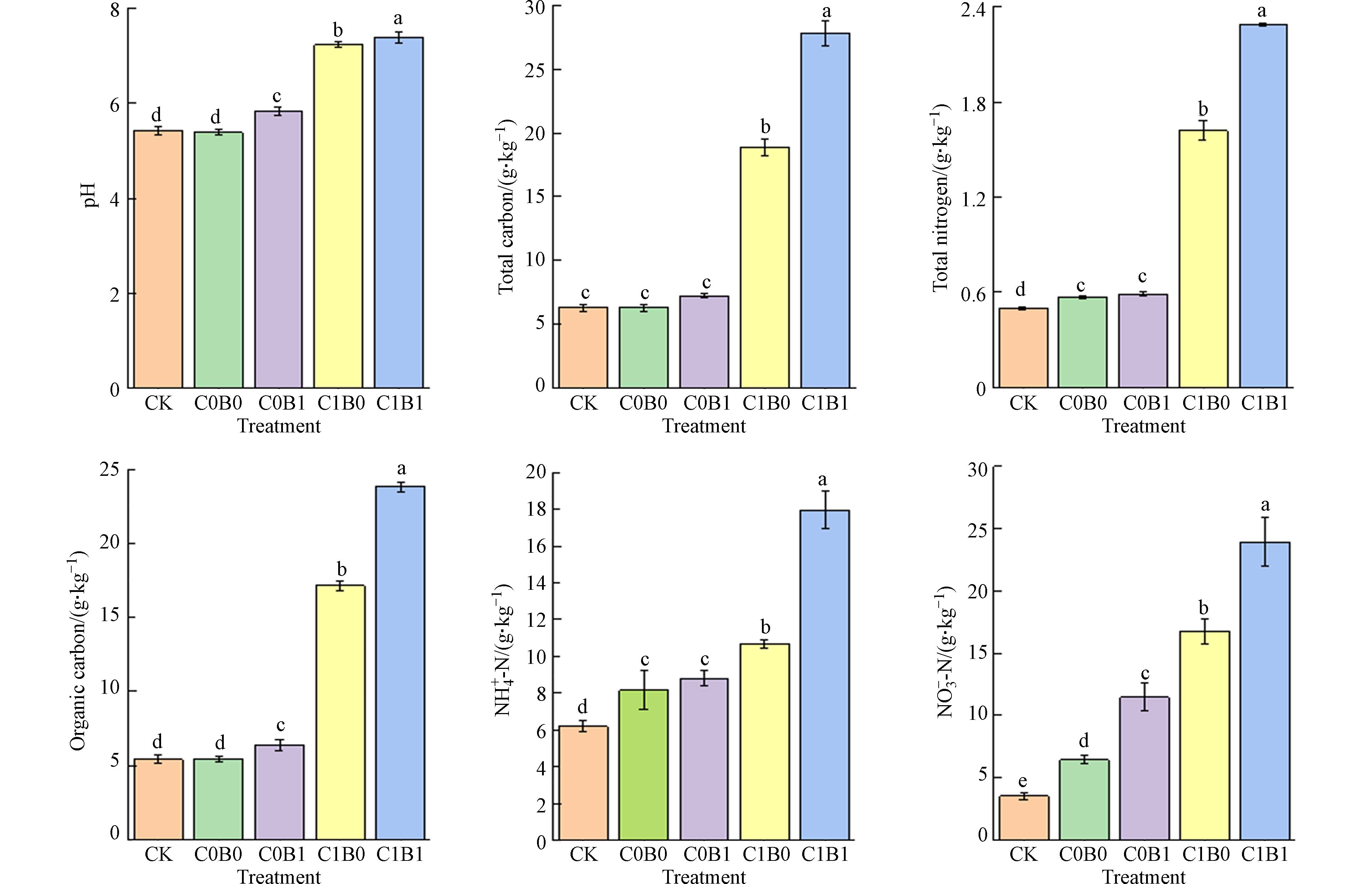 图 1 不同配施条件下土壤的理化性质Figure 1. Physical and chemical properties of soil under different application conditions同组不同字母在同组中表示有显著差异(P<0.05)Different letters in the same group meant significant difference at 0.05 level.
图 1 不同配施条件下土壤的理化性质Figure 1. Physical and chemical properties of soil under different application conditions同组不同字母在同组中表示有显著差异(P<0.05)Different letters in the same group meant significant difference at 0.05 level.2.2 生物质炭与沼液施用对土壤碳固定与氮代谢途径微生物群落丰度的影响
通过对土壤微生物群落丰度的检测,得到不同处理下土壤细菌丰度情况,将相对丰度超过1%的微生物定义为优势菌. 从图2a可以看出,5个处理的优势菌门相同,主要为:变形菌门(Proteobacteria)、绿弯菌门(Chloroflexi)、放线菌门(Actinobacteria)、酸杆菌门(Acidobacteria)、浮霉菌门(Planctomycetes)、芽单胞菌门(Gemmatimonadetes)、蓝光合菌门(Cyanobacteria)和装甲菌门(Armatimonadetes)共8个,占总菌门91.8%—97.7%. 相比于CK处理,C0B0处理显著增加Cyanobacteria的相对丰度,而显著降低Proteobacteria、Gemmatimonadetes和Armatimonadetes的相对丰度,C0B1、C1B0和C1B1处理显著增加Chloroflexi、Actinobacteria和Acidobacteria的相对丰度,显著降低Cyanobacteria与Armatimonadetes的相对丰度.
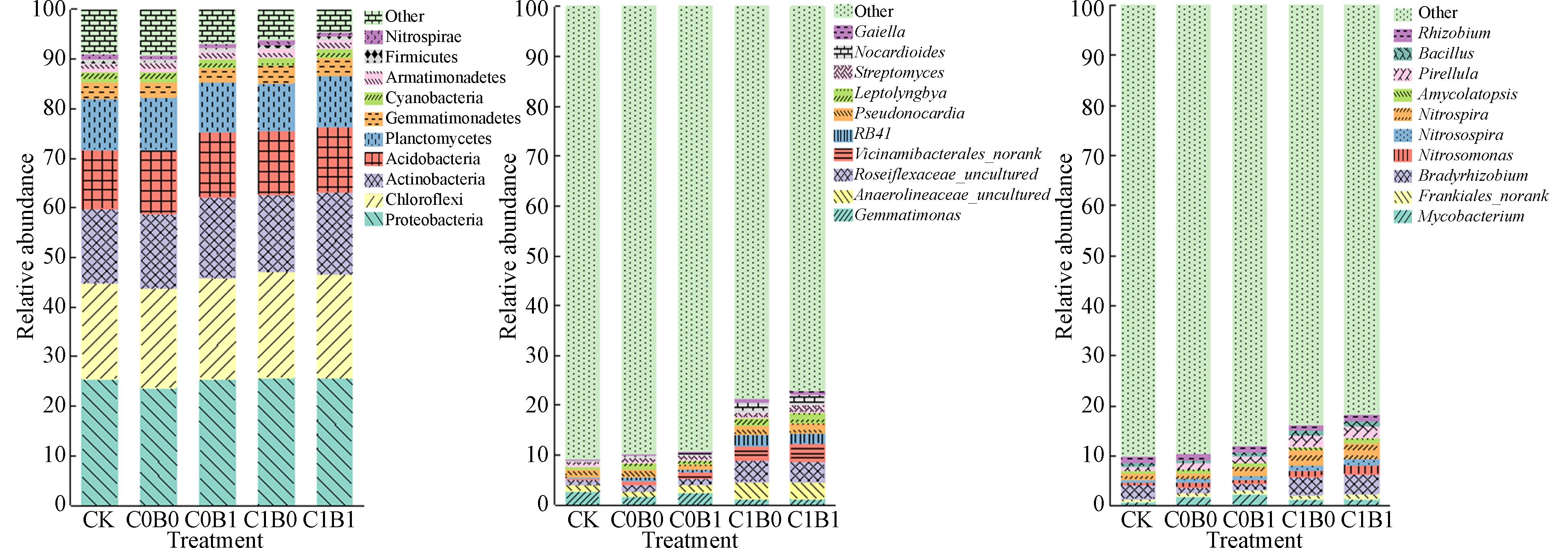 图 2 不同处理下门水平与属水平上相对丰度情况Figure 2. Relative abundance of different components at the phylum level and genus level(a)不同处理门水平相对丰度;(b)不同处理属水平碳固定菌属相对丰度;(c)不同处理属水平氮代谢菌属相对丰度(a) relative abundance of phylum levels in different treatments;(b) relative abundance of carbon-fixing bacteria in different treatments; (c) relative abundance of nitrogen metabolizing bacteria in different treatments
图 2 不同处理下门水平与属水平上相对丰度情况Figure 2. Relative abundance of different components at the phylum level and genus level(a)不同处理门水平相对丰度;(b)不同处理属水平碳固定菌属相对丰度;(c)不同处理属水平氮代谢菌属相对丰度(a) relative abundance of phylum levels in different treatments;(b) relative abundance of carbon-fixing bacteria in different treatments; (c) relative abundance of nitrogen metabolizing bacteria in different treatments由图2b可知,各处理中碳固定优势菌属为Gemmatimonas、未分类的厌氧绳菌属(Anaerolineaceae_uncultured)、未分类的绿弯菌属(Roseiflexaceae_uncultured)、假诺卡氏菌属(Pseudonocardia)和Streptomyces. 且不同处理间碳固定优势菌属丰度差异显著,其中碳固定优势菌属丰度在C1B1中达最高,相比于CK处理,增涨64%. 与CK处理相比较,单独施加化肥、生物质炭和炭-沼液联合施用(C1B1)的处理中Gemmatimonas丰度显著降低,分别降低了39.0%、55.4%和51.3%. 生物质炭的单施或与沼液混施对Anaerolineaceae_uncultured和Roseiflexaceae_uncultured菌属的相对丰度有显著提高,单施生物质炭时提高168.7%和156.1%,生物质炭和沼液联合施用时提高246.4%和220.2%. Pseudonocardia菌属在单施沼液处理中显著降低18.2%,施加生物质炭的处理中则显著增加,其中C1B0与C1B1分别增加43.7%和50.3%. C1B1中Streptomyces菌属丰度最高,相比于CK与C0B0处理,分别提高74.0%和65.6%,这可能有两方面原因;一方面是生物质炭通过吸附沼液中的养分,储存于生物质炭的孔隙中,缓解沼液在高温条件下的挥发损失,提供了充足的养料,促进土壤中的菌群丰度的增加. 另一方面可能是生物质炭的施用导致土壤pH值升高,使得菌群的生存环境得到改善,再联合沼液施用,增加土壤养分含量,提高土壤支持多种生态系统功能的能力,最终导致菌群丰度的显著提升[38].
由图2c可知,分枝杆菌属(Mycobacterium)、Bradyrhizobium、Nitrospira、小梨形菌属(Pirellula)和根瘤菌(Rhizobium)为氮代谢优势菌属,且优势菌属丰度在不同处理间差异达显著水平,且生物质炭与沼液联合施用处理中丰度达到最高值,相比于CK处理,增涨65.7%. 施用化肥、沼液或生物质炭的处理中Mycobacterium菌属丰度相比于CK处理均有显著提升,其中沼液单独施用提升最显著,达到139.4%. 此外,单施沼液处理中Rhizobium菌属丰度比单施化肥处理增加了20.8%,这可能是由于生物质炭的施用刺激了固氮菌的活性,并且通过利用沼液中丰富的养分实现丰度的增加[39 − 40]. Bradyrhizobium菌属丰度在生物质炭单施或与沼液配施时显著增加35.1%和41.8%,而单施化肥或沼液的处理中显著降低了57.2%和51.8%. 生物质炭和沼液配施的处理中Nitrospira和Pirellula菌群丰度显著高于其他处理,相比于CK处理显著提升148.7%和115.7%,这一方面可能是由于生物质炭的多孔隙结构增加了土壤的透气性,提高了土壤中氧气含量,引起好氧菌丰度的增加[41]. 另一方面可能是由于沼液施用引入了丰富的氮素养分,为硝化作用提供了充足的原料[42]. 由此说明,长期使用生物质炭与沼液,有利于维护土壤微生态环境的健康稳定,对土壤碳氮循环产生有利影响.
2.3 不同施肥模式对土壤碳固定与氮代谢功能基因的影响
利用荧光定量RCR法对土壤功能基因测定,注释到13种碳固定基因和22种氮代谢基因. 对比基因测试结果,选取基因丰度排名前10的功能基因进行组间分析. 通过碳固定基因结果可知(图3a),相较于CK与C0B0处理,C1B1的中的基因相对丰度显著增加,其中acsE相对丰度分别提高102.2%与97.4%,rbcL基因分别提高175.8%和95.3%. 沼液或生物质炭的单独施用,对accA、acsA、acsE、rbcL、cdaR、korA、frdA、pccA、acsB的基因相对丰度显著高于单施化肥,其中C0B1和C1B0中acsE的丰度分别提高35.4%与47.8%,这一方面可能是由于生物质炭自身的难降解性导致土壤碳库的增加,间接有利于土壤的固碳[43]. 另一方面,沼液施用提高了土壤中养分浓度,改变了微生物的代谢途径[44]. 对氮代谢基因检测结果进行组间分析(图3b),其中硝化型功能基因为amoB、amoA2、nxrA,反硝化型功能基因为narG、napA、nirK3、nosZ2、nirS1,氮矿化型功能基因gdhA,固氮型功能基因nifH. 与CK相比较,C0B0、C0B1、C1B0和C1B1处理nxrA、narG、napA、nirK3与gdhA功能基因丰度显著提高,这可能是由于生物质炭与沼液施用,引入了大量的活性微生物,创建了一个富营养化的微环境领域,补充了硝化细菌、反硝化细菌和矿化细菌所需的底物,刺激了氮代谢细菌的繁殖,导致基因丰度的增加[45]. 与C0B0处理相比较,C0B1处理nifH、nxrA、nirK3基因相对丰度显著增加,C1B0处理中nxrA、narG、napA、nirS1基因丰度有显著增加,C1B1处理中硝化型基因nxrA显著增加54.0%,反硝化基因narG、napA、nirK3、nirS1也显著增加,这与郑一玲等[46]研究一致. 这可能是由于外部碳源与氮源的加入,为土壤微生物提供富足的养分环境,使得微生物通过调整基因表达来适应环境变化,进而提高功能基因的表达[47]. 总体分析可知,长期生物质炭与沼液替代化肥施用,可促进土壤碳固定与氮代谢途径基因的表达,提升土壤微生物参与碳氮循环潜力和代谢特征.
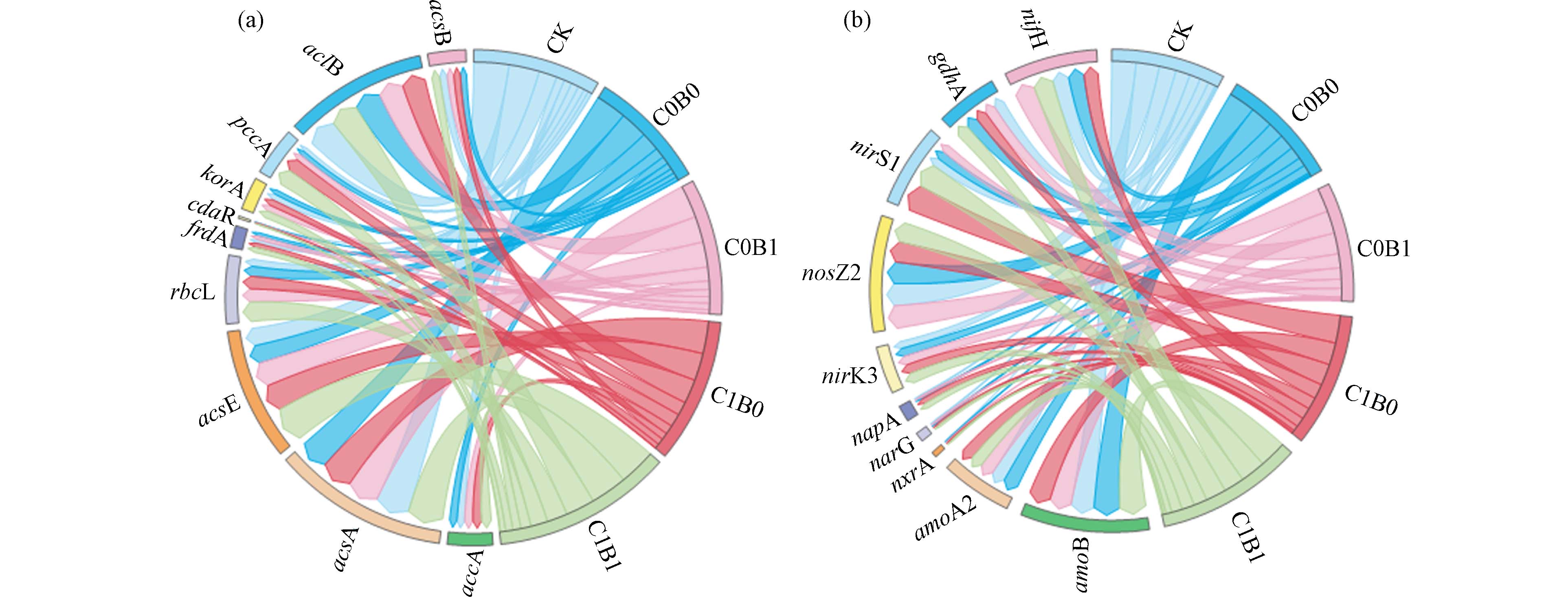 图 3 碳固定与氮代谢关键基因的组间比较Figure 3. Inter group comparison of key genes involved in carbon fixation and nitrogen metabolism(a)不同处理碳固定功能基因相对丰度;(b)不同处理氮代谢功能基因相对丰度(a) Relative abundance of functional genes for carbon fixation under different treatments; (b) relative abundance of functional genes for nitrogen metabolism under different treatments
图 3 碳固定与氮代谢关键基因的组间比较Figure 3. Inter group comparison of key genes involved in carbon fixation and nitrogen metabolism(a)不同处理碳固定功能基因相对丰度;(b)不同处理氮代谢功能基因相对丰度(a) Relative abundance of functional genes for carbon fixation under different treatments; (b) relative abundance of functional genes for nitrogen metabolism under different treatments2.4 碳固定与氮代谢途径相关微生物与功能基因的相关性分析
为了解碳固定与氮代谢微生物的生物学特性与生态学适应性,选取丰度排名前十的微生物(属水平)及关键型功能基因进行相关性分析. 由图4a可知,碳固定功能基因与微生物之间正相关性(85%)的占比大于负相关性(15%),10种菌属中Vicinamibacterales_norank、Streptomyces和Gaiella与功能基因如accA、acsE、frdA、cdaR和pccA呈显著正相关,表明3种细菌在碳固定途径中起到重要的固碳作用. 此外rbcL基因与Vicinamibacterales_norank和Streptomyces菌属也存在显著正相关. 并且frdA和pccA基因与菌属Anaerolineaceae_uncultured、Roseiflexaceae_uncultured、Pseudonocardia和Nocardioides呈显著正相关. 基因pccA与RB41呈显著正相关.
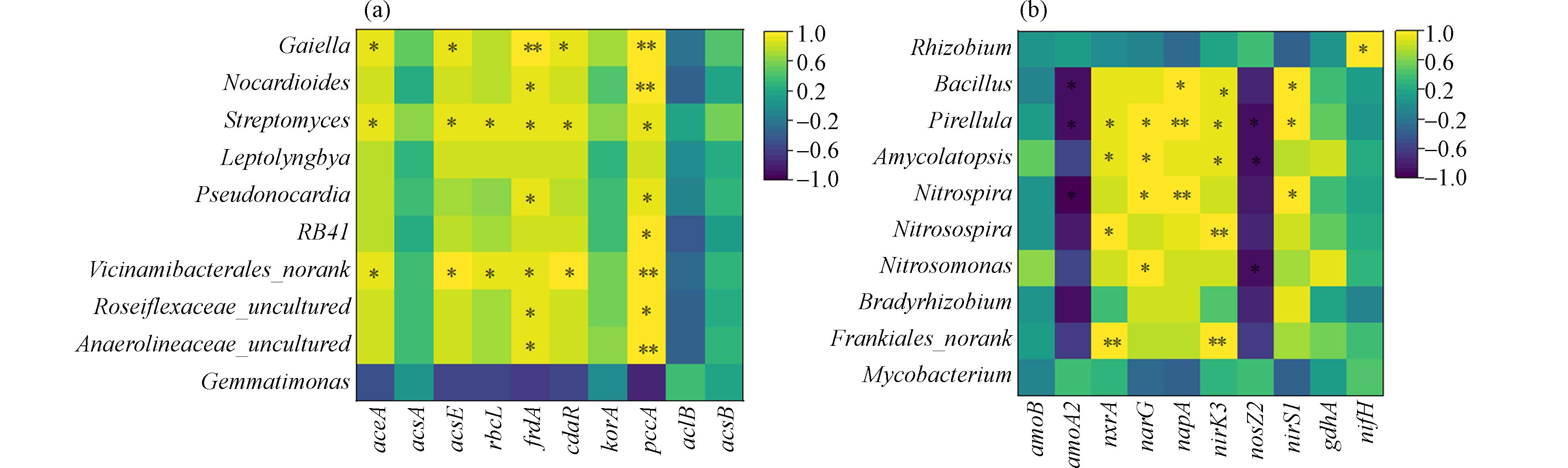 图 4 功能基因与微生物菌属相关性分析Figure 4. Correlation analysis between functional genes and microbial genera(a)碳固定优势菌属与相关功能基因的相关性;(b)氮代谢优势菌属与相关功能基因的相关性(a) Correlation between carbon-fixed dominant bacteria and related functional genes; (b) Correlation between nitrogen metabolism dominant bacteria and related functional genes *P<0.05,**P<0.01
图 4 功能基因与微生物菌属相关性分析Figure 4. Correlation analysis between functional genes and microbial genera(a)碳固定优势菌属与相关功能基因的相关性;(b)氮代谢优势菌属与相关功能基因的相关性(a) Correlation between carbon-fixed dominant bacteria and related functional genes; (b) Correlation between nitrogen metabolism dominant bacteria and related functional genes *P<0.05,**P<0.01由图4b可知,氮代谢关键基因与主要微生物之间正相关性(73%)的占比大于负相关性(27%). 其中硝化型基因(nxrA)和反硝型基因(nirK3)与Frankiales_norank、Nitrosospira、Amycolatopsis和Pirellula菌属之间显著正相关. 反硝化基因(napA和nirS1)与Nitrospira、Pirellula和Bacillus之间显著正相关. Nitrosomonas、Nitrospira、Amycolatopsis和Pirellula4种菌属与反硝化型基因(narG)显著正相关. Rhizobium与氮固定型基因(nifH)显著正相关. Bacillus菌属与nirK3显著正相关,而与有氧氨氧化型基因(amoA2)显著负相关. 此外,amoA2还与Pirellula和Nitrospira显著负相关. 反硝化型基因(nosZ2)与Pirellula、Amycolatopsis和Nitrosomonas3种菌属显著负相关.
3. 结论(Conclusion)
采取生物质炭与沼液联合施用可显著增加土壤有机碳、全碳、全氮、铵态氮、硝态氮含量并提高土壤pH. 有益于碳固定与氮代谢微生物属水平种群丰度的增加,可以提高固碳功能基因与氮代谢功能基因表达水平,增强土壤微生物的固碳与反硝化能力,促进土壤系统的碳、氮循环. 因此采取生物质炭与沼液联合替代化肥施用,有利于维护土壤微环境健康的同时增强微生物的生态适应性,保障土地生产力的稳定.
-
图 1 不同配施条件下土壤的理化性质
Figure 1. Physical and chemical properties of soil under different application conditions
图 2 不同处理下门水平与属水平上相对丰度情况
Figure 2. Relative abundance of different components at the phylum level and genus level
图 3 碳固定与氮代谢关键基因的组间比较
Figure 3. Inter group comparison of key genes involved in carbon fixation and nitrogen metabolism
-
[1] 刘晏君, 吕卫光, 白娜玲, 等. 稻鳝生态种养不同施肥处理对土壤理化及微生物性状的影响[J]. 土壤通报, 2023, 54(6): 1384-1391. LIU Y J, LV W G, BAI N L, et al. Effects of different fertilization treatments of ecological rice-eel co-culture on soil physicochemical and microbial properties[J]. Chinese Journal of Soil Science, 2023, 54(6): 1384-1391 (in Chinese).
[2] 李文博, 刘少君, 叶新新, 等. 稻田综合种养模式对土壤生态系统的影响研究进展[J]. 生态与农村环境学报, 2021, 37(10): 1292-1300. LI W B, LIU S J, YE X X, et al. Effects of the co-culture of rice and aquatic animals on soil eco-system: A review[J]. Journal of Ecology and Rural Environment, 2021, 37(10): 1292-1300 (in Chinese).
[3] SHI C, ZHANG Q W, YU B W. Higher improvement in soil health by animal-sourced than plant-sourced organic materials through optimized substitution[J]. Agriculture, Ecosystems & Environment, 2024, 363: 108875. [4] 杜彩艳, 鲁海燕, 熊艳竹, 等. 连续两年沼液与化肥配施对桃生长及土壤理化性质的影响[J]. 中国农业科技导报, 2023, 25(8): 165-175. DU C Y, LU H Y, XIONG Y Z, et al. Effects of combined application of biogas slurry and chemical fertilizer on peach growth and soil physical and chemical properties for two consecutive years[J]. Journal of Agricultural Science and Technology, 2023, 25(8): 165-175 (in Chinese).
[5] 柴彦君, 张睿, 江建锋, 等. 沼液化肥配施对芦笋地土壤肥力及芦笋品质的影响[J]. 农业工程学报, 2023, 39(5): 120-127. CHAI Y J, ZHANG R, JIANG J F, et al. Effects of the combined biogas slurry with chemical fertilizer on soil fertility and Asparagus quality in field[J]. Transactions of the Chinese Society of Agricultural Engineering, 2023, 39(5): 120-127 (in Chinese).
[6] 金柯达, 王绍轩, 胡宝娥, 等. 沼肥施用量对上海青产量及土壤理化特性的影响[J]. 中国沼气, 2022, 40(6): 50-56. JIN K D, WANG S X, HU B E, et al. Effects of biogas fertilizer application rate on Shanghai cabbage yield and soil physicochemical properties[J]. China Biogas, 2022, 40(6): 50-56 (in Chinese).
[7] 滕云飞, 尚斌, 陶秀萍. 猪粪沼液对设施基质栽培番茄的营养效应[J]. 中国农业科学, 2023, 56(19): 3869-3878. TENG Y F, SHANG B, TAO X P. Nutritional effects of liquid digestate on tomatoes grown in facility substrates[J]. Scientia Agricultura Sinica, 2023, 56(19): 3869-3878 (in Chinese).
[8] 刘强, 袁延飞, 刘一帆, 等. 生物炭对盐渍化土壤改良的研究进展[J]. 地球科学进展, 2022, 37(10): 1005-1024. doi: 10.11867/j.issn.1001-8166.2022.050 LIU Q, YUAN Y F, LIU Y F, et al. Research progress: The application of biochar in the remediation of salt-affected soils[J]. Advances in Earth Science, 2022, 37(10): 1005-1024 (in Chinese). doi: 10.11867/j.issn.1001-8166.2022.050
[9] 刘红梅, 李睿颖, 高晶晶, 等. 保护性耕作对土壤团聚体及微生物学特性的影响研究进展[J]. 生态环境学报, 2020, 29(6): 1277-1284. LIU H M, LI R Y, GAO J J, et al. Research progress on the effects of conservation tillage on soil aggregates and microbiological characteristics[J]. Ecology and Environmental Sciences, 2020, 29(6): 1277-1284 (in Chinese).
[10] XIA L L, LAM S K, CHEN D L, et al. Can knowledge-based N management produce more staple grain with lower greenhouse gas emission and reactive nitrogen pollution?A meta-analysis[J]. Global Change Biology, 2017, 23(5): 1917-1925. doi: 10.1111/gcb.13455 [11] FANG H, WANG H F, CAI L, et al. Prevalence of antibiotic resistance genes and bacterial pathogens in long-term manured greenhouse soils as revealed by metagenomic survey[J]. Environmental Science & Technology, 2015, 49(2): 1095-1104. [12] LIU S W, ZHANG Y J, ZONG Y J, et al. Response of soil carbon dioxide fluxes, soil organic carbon and microbial biomass carbon to biochar amendment: A meta-analysis[J]. GCB Bioenergy, 2016, 8(2): 392-406. doi: 10.1111/gcbb.12265 [13] 刘茗, 曹林桦, 刘彩霞, 等. 亚热带4种典型森林植被土壤固碳细菌群落结构及数量特征[J]. 土壤学报, 2021, 58(4): 1028-1039. LIU M, CAO L H, LIU C X, et al. Characterization of population and community structure of carbon-sequestration bacteria in soils under four types of forest vegetations typical of subtropical zone[J]. Acta Pedologica Sinica, 2021, 58(4): 1028-1039 (in Chinese).
[14] 杨玲, 张艺, 钟俊杰, 等. 不同调酸剂对种植玉米红壤微生物群落的影响[J]. 农业环境科学学报, 2024, 43(3): 609-616. YANG L, ZHANG Y, ZHONG J J, et al. Effects of different acid modulators on the microbial communities in maize planting red soil[J]. Journal of Agro-Environment Science, 2024, 43(3): 609-616 (in Chinese).
[15] 张家鹏, 李为萍, 池曌男, 等. 地下滴灌对向日葵根区土壤微环境及细菌氮转化功能基因的影响[J]. 环境科学学报, 2024, 44(5): 474-485. ZHANG J P, LI W P, CHI Z N, et al. Effects of subsurface drip irrigation on soil microenvironment and functional genes for bacterial nitrogen transformation in sunflower root zone[J]. Acta Scientiae Circumstantiae, 2024, 44(5): 474-485 (in Chinese).
[16] 刘倩, 陈晓, 李彦澄, 等. 湖泊沉积物的氮代谢微生物、功能基因及代谢途径分析: 以贵州省红枫湖为例[J]. 环境科学, 2024, 45(10): 6086-6095. LIU Q, CHEN X, LI Y C, et al. Analysis of nitrogen metabolism microorganisms, functional genes and metabolic pathways in lake sediments: A case study of Hongfeng Lake in Guizhou Province[J]. Environmental Science, 2024, 45(10): 6086-6095 (in Chinese).
[17] 韩磊, 胡盎, 任明磊, 等. 湖泊微生物群落及其介导的碳循环过程[J]. 生命科学, 2023, 35(12): 1613-1629. HAN L, HU A, REN M L, et al. Lake microbial communities and their mediated carbon cycling processes[J]. Chinese Bulletin of Life Sciences, 2023, 35(12): 1613-1629 (in Chinese).
[18] 王小利, 王淑兰. 基于KEGG的碳固定和氮代谢通路土壤微生物组筛选[J]. 农业机械学报, 2020, 51(8): 303-310. WANG X L, WANG S L. Soil microbiome screening for carbon fixation and nitrogen metabolism pathways based on KEGG database[J]. Transactions of the Chinese Society for Agricultural Machinery, 2020, 51(8): 303-310 (in Chinese).
[19] 李凯凯, 张丙昌, 赵康, 等. 毛乌素沙地固碳功能菌群落随生物结皮发育阶段的演变特征[J]. 生态学报, 2024, 44(3): 1177-1190. LI K K, ZHANG B C, ZHAO K, et al. Succession of carbon-fixing microbial community in different stages of biological soil crusts in the Mu Us Sandy Land[J]. Acta Ecologica Sinica, 2024, 44(3): 1177-1190 (in Chinese).
[20] 宋达成, 赵文智, 李广宇, 等. 退耕对民勤绿洲土壤碳氮循环关键微生物及功能基因的影响[J]. 生态学报, 2024, 44(2): 805-818. SONG D C, ZHAO W Z, LI G Y, et al. Effects of abandoned farmland on key microorganisms and functional genes of soil carbon and nitrogen cycles in Minqin Oasis[J]. Acta Ecologica Sinica, 2024, 44(2): 805-818 (in Chinese).
[21] 王彬浩, 吴愉萍, 陈彦博, 等. 秸秆还田对土壤微生物功能基因和功能类群的影响[J]. 浙江大学学报(农业与生命科学版), DOI:10.3785/j.issn.1008-9209.2023.07.172. WANG B H, WU Y P, CHEN Y B, et al. Effects of straw returning on soil microbial functional genes and functional groups[J]. Journal of ZheJiang University (Agriculture and Life Sciences), DOI:10.3785/j.issn.1008-9209.2023.07.172 (in Chinese).
[22] 舒洋, 陈金平, 丁兆华, 等. 林火强度对兴安落叶松林土壤氮循环功能基因的影响[J]. 草地学报, 2024, 32(3): 726-735. SHU Y, CHEN J P, DING Z H, et al. Effects of forest fire intensity on functional genes of soil nitrogen cycling in Larix gmelinii[J]. Acta Agrestia Sinica, 2024, 32(3): 726-735 (in Chinese).
[23] 王宇峰, 孟会生, 李廷亮, 等. 培肥措施对复垦土壤微生物碳氮代谢功能多样性的影响[J]. 农业工程学报, 2020, 36(24): 81-90. WANG Y F, MENG H S, LI T L, et al. Effects of fertilization regime on the functional diversity of microbial carbon and nitrogen metabolism in reclaimed soil[J]. Transactions of the Chinese Society of Agricultural Engineering, 2020, 36(24): 81-90 (in Chinese).
[24] 马慧霞, 张桥, 陈会巧, 等. 长期有机培肥对南方红壤区稻田土壤碳循环功能基因的影响[J]. 南方农业学报, 2023, 54(5): 1405-1416. MA H X, ZHANG Q, CHEN H Q, et al. Effects of long-term organic fertilization on the functional genes of carbon cycle in paddy soils[J]. Journal of Southern Agriculture, 2023, 54(5): 1405-1416 (in Chinese).
[25] 赵姣, 马静, 朱燕峰, 等. 植被类型对黄土高原露采矿山复垦土壤碳循环功能基因的影响[J]. 环境科学, 2023, 44(6): 3386-3395. ZHAO J, MA J, ZHU Y F, et al. Effects of vegetation types on carbon cycle functional genes in reclaimed soil from open pit mines in the Loess Plateau[J]. Environmental Science, 2023, 44(6): 3386-3395 (in Chinese).
[26] PENG S M, LIU W, XU G, et al. A meta-analysis of soil microbial and physicochemical properties following native forest conversion[J]. CATENA, 2021, 204: 105447. doi: 10.1016/j.catena.2021.105447 [27] LI H, YE D D, WANG X G, et al. Soil bacterial communities of different natural forest types in Northeast China[J]. Plant and Soil, 2014, 383(1): 203-216. [28] CHEN J, SINSABAUGH R L. Linking microbial functional gene abundance and soil extracellular enzyme activity: Implications for soil carbon dynamics[J]. Global Change Biology, 2021, 27(7): 1322-1325. doi: 10.1111/gcb.15506 [29] ZHAO M X, XUE K, WANG F, et al. Microbial mediation of biogeochemical cycles revealed by simulation of global changes with soil transplant and cropping[J]. The ISME Journal, 2014, 8(10): 2045-2055. doi: 10.1038/ismej.2014.46 [30] 魏甲彬, 李有清. 生物炭添加对根际土壤微生物群落影响研究进展[J]. 湖南生态科学学报, 2023, 10(2): 101-108. WEI J B, LI Y Q. Research progress on the effects of biochar addition on rhizosphere soil microbial communities[J]. Journal of Hunan Ecological Science, 2023, 10(2): 101-108 (in Chinese).
[31] 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科学技术出版社, 2000. LU R K. Methods of soil agrochemical analysis[M]. China Agricultural Science and Technology Press, 2000(in Chinese).
[32] ZHENG B X, ZHU Y G, SARDANS J, et al. QMEC: A tool for high-throughput quantitative assessment of microbial functional potential in C, N, P, and S biogeochemical cycling[J]. Science China Life Sciences, 2018, 61(12): 1451-1462. doi: 10.1007/s11427-018-9364-7 [33] CHEN Q L, DING J, LI C Y, et al. Microbial functional attributes, rather than taxonomic attributes, drive top soil respiration, nitrification and denitrification processes[J]. Science of the Total Environment, 2020, 734: 139479. doi: 10.1016/j.scitotenv.2020.139479 [34] 覃潇敏, 潘浩男, 环秀菊, 等. 生物炭对菠萝连作土壤微生物学特性及其心腐病的影响[J]. 中国土壤与肥料, 2024(2): 65-71. QIN X M, PAN H N, HUAN X J, et al. Effects of biochar on soil microbial characteristics of pineapple and the heart rot under continuous cropping[J]. Soil and Fertilizer Sciences in China, 2024(2): 65-71 (in Chinese).
[35] 王强, 耿增超, 许晨阳, 等. 施用生物炭对塿土土壤微生物代谢养分限制和碳利用效率的影响[J]. 环境科学, 2020, 41(5): 2425-2433. WANG Q, GENG Z C, XU C Y, et al. Effects of biochar application on soil microbial nutrient limitations and carbon use efficiency in Lou soil[J]. Environmental Science, 2020, 41(5): 2425-2433 (in Chinese).
[36] 温云杰, 张纪涛, 李琳, 等. 沼液配施化肥对大葱产量和土壤养分、微生物及酶活性的影响[J]. 中国生态农业学报(中英文), 2024, 32(1): 95-105. WEN Y J, ZHANG J T, LI L, et al. Effects of biogas slurry combined with chemical fertilizer on Allium fistulosum yields, soil nutrients, microorganisms, and enzymes activities[J]. Chinese Journal of Eco-Agriculture, 2024, 32(1): 95-105 (in Chinese).
[37] LIANG P Y, YU H O, HUANG J L, et al. The review on adsorption and removing ammonia nitrogen with biochar on its mechanism[J]. MATEC Web of Conferences, 2016, 67: 07006. doi: 10.1051/matecconf/20166707006 [38] 杨雪, 曹霞, 白冰, 等. 根施生物炭对设施连作土壤氮素转化及黄瓜幼苗根系氮代谢的影响[J]. 应用生态学报, 2024, 35(3): 713-720. YANG X, CAO X, BAI B, et al. Effects of root-applied biochar on soil nitrogen transformation and root nitrogen metabolism of cucumber seedlings in facility continuous cropping soils[J]. Chinese Journal of Applied Ecology, 2024, 35(3): 713-720 (in Chinese).
[39] 姚玲丹, 程广焕, 王丽晓, 等. 施用生物炭对土壤微生物的影响[J]. 环境化学, 2015, 34(4): 697-704. YAO L D, CHENG G H, WANG L X, et al. Effects of biochar application to microorganisms in soil[J]. Environmental Chemistry, 2015, 34(4): 697-704 (in Chinese).
[40] 李钰飞, 许俊香, 刘本生, 等. 不同来源沼液对土壤微生物群落碳代谢的影响[J]. 中国生态农业学报(中英文), 2021, 29(11): 1921-1930. LI Y F, XU J X, LIU B S, et al. Effects of different biogas slurries on soil microbial carbon metabolism[J]. Chinese Journal of Eco-Agriculture, 2021, 29(11): 1921-1930 (in Chinese).
[41] 陈健, 卢伟伟, 杜娅茹, 等. 生物质炭和氮肥对马尾松人工林土壤微生物群落结构和酶活性的影响[J]. 环境科学学报, 2024, 44(4): 333-343. CHEN J, LU W W, DU Y R, et al. Effects of biochar and nitrogen fertilizer on soil microbial community structure and enzyme activity in a Pinus massoniana plantation[J]. Acta Scientiae Circumstantiae, 2024, 44(4): 333-343 (in Chinese).
[42] 唐存柳, 靳泽文, 姚光伟, 等. 生物质炭-沼液联合施用对调控氮循环功能基因促进氮素增效的影响[J]. 环境化学, 2023, 42(8): 2843-2852. doi: 10.7524/j.issn.0254-6108.2015.04.2014072802 TANG C L, JIN Z W, YAO G W, et al. Effects of combined application of biochar and biogas slurry on regulating nitrogen cycle function genes and promoting nitrogen efficiency[J]. Environmental Chemistry, 2023, 42(8): 2843-2852 (in Chinese). doi: 10.7524/j.issn.0254-6108.2015.04.2014072802
[43] LU W W, DING W X, ZHANG J H, et al. Biochar suppressed the decomposition of organic carbon in a cultivated sandy loam soil: A negative priming effect[J]. Soil Biology and Biochemistry, 2014, 76: 12-21. doi: 10.1016/j.soilbio.2014.04.029 [44] 刘银秀, 池永清, 董越勇, 等. 不同沼液施用年限土壤养分含量和微生物群落结构差异[J]. 植物营养与肥料学报, 2023, 29(3): 483-495. LIU Y X, CHI Y Q, DONG Y Y, et al. Variation of nutrient content and microbial community in soils under different application years of biogas slurry[J]. Journal of Plant Nutrition and Fertilizers, 2023, 29(3): 483-495 (in Chinese).
[45] 陈会巧, 马慧霞, 张桥, 等. 长期培肥降低稻田土壤硝化和反硝化细菌功能基因丰度并减缓氮素周转[J]. 植物营养与肥料学报, 2023, 29(9): 1630-1642. doi: 10.11674/zwyf.2023098 CHEN H Q, MA H X, ZHANG Q, et al. Long-term fertilization reduces nitrifying and denitrifying functional gene abundance and slows down the nitrogen recycle in paddy soils[J]. Journal of Plant Nutrition and Fertilizers, 2023, 29(9): 1630-1642 (in Chinese). doi: 10.11674/zwyf.2023098
[46] 郑一玲, 范栩妙, 钟哲伦, 等. 生物质炭-沼液配施条件下旱地红壤碳氮循环功能基因丰度主控因子与耦合关系[J]. 农业资源与环境学报, 2024, 41(4): 835-845. ZHENG Y L, FAN X M, ZHONG Z L, et al. The main controlling factors and coupling relationship of functional gene abundance in carbon and nitrogen cycling in dryland red soil with biochar and biogas slurry co-application[J]. Journal of Agricultural Resources and Environment, 2024, 41(4): 835-845 (in Chinese).
[47] 何瑞鹏, 徐婉茹, 刘一萌, 等. 若尔盖沙化土地治理土壤微生物群落与功能基因比较研究[J]. 微生物学报, 2024, 64(6): 2071-2090. HE R P, XU W R, LIU Y M, et al. Comparison of soil microbial communities and functional genes in sandy land management of Zoige[J]. Acta Microbiologica Sinica, 2024, 64(6): 2071-2090 (in Chinese).
-

 点击查看大图
点击查看大图
计量
- 文章访问数: 580
- HTML全文浏览数: 580
- PDF下载数: 7
- 施引文献: 0
