

-
水体富营养化(Eutrophication)是由于水生系统的氮、磷等营养元素的含量过高、相对比例失调或化学形态变化,引起的水体功能失衡的现象[1],会导致水中浮游植物、浮游动物等群落结构变化,水质迅速恶化[2]。随着城市化和工业化的进行,大量的含氮物质随生活污水、工农业废水等进入水体并积累,对水生生态环境和人体健康构成威胁[3-6]。针对富营养水体,通常采用生物法进行治理:放养田螺、河蚬等滤食性动物可摄食水中藻类,提高水体透明度,有利于沉水植物与微生物增殖,从而降低氮浓度,但需要考虑水生动物对富营养水环境的适应性问题[7-8];种植水生植物可吸收、利用水体中氮源促进脱氮,但在植物成熟后效果较差[9-12];微生物脱氮可持续通过氨化、硝化-反硝化、厌氧氨氧化等作用使氮进行形态转化,并把氮转化为气态或固定在生物体内而脱离水体[13-15],相较于动物法和植物法,微生物对环境的适应性强且能持续发挥其作用。
碳氮比(C/N)是微生物脱氮的重要影响因素[16-18],在适宜范围内,随着C/N的升高,异化硝酸盐还原的功能基因和反硝化过程相关酶的相对丰度均增大[19],进而促进氮的去除。BONASSA等[20]认为传统的硝化/反硝化工艺需要高于3.5的CODb/N以实现完全的脱氮,其中CODb是由生物降解导致的COD去除量。CHIU等[21]认为支持常规生物反硝化的最佳COD/
$ {\text{NO}}_{\text{3}}^{{-}}\text{-N} $ 比率为6.0~8.0。但国内外污废水普遍存在C/N低的问题使生物脱氮受到抑制[22-25]。一些研究采用投加甲醇、乙醇、乙酸钠、葡萄糖等可溶有机碳源的方式对水体C/N进行调节以促进脱氮,但该方式成本高、易产生二次污染,且对投加技术要求高[26-27] 。中国种植业每年可产生数亿吨生物质固体废弃物,这些生物质固废的成分安全无害,在水中能持续释放有机碳源,且能增大水体中的固-液界面面积,有利于微生物附着生长[28-30]和水体氮的转化与去除[30-31]。以生物质代替可溶性碳源,可降低碳源材料的购买、运输和存储成本,减弱碳源投加的操作难度,降低碳源添加频率[32-34]。雷柯柯等[35]分别以甘蔗渣、稻壳粉为载体,使水体絮团总菌数量提高至105~107 cfu·L−1(cfu为活菌培养计数时的菌落形成单位),罗佳等[36]向富营养化水体中添加稻壳、木屑,水体中硝化细菌、反硝化细菌数量和细菌多样性也高于对照组。将15 g玉米芯、花生壳、稻壳在3 L超纯水中浸泡,10 d后COD释放量分别达到了225.88、178.78、384.16 mg·L−1,在静态反硝化实验中三者的
$ {\text{NO}}_{\text{3}}^{-} $ 去除率均在3 d内达到了85%以上[37]。将稻草、丝瓜络、玉米芯分别加入生态袋中进行实验,$ {\text{NH}}_{\text{4}}^{\text{+}}\text{-N} $ 平均去除率分别为78.4%、77.8%和73.6%,TN去除率达82.7%、79.6%、77.4%,明显高于对照组的68.1%和71.7%[38]。生物质粒径决定了生物质与水体的固-液接触面积的大小,这不仅会影响生物质与周围水体的物质交换通量[39],也可能导致脱氮效果的差异。目前工程和实验中选取特定粒径的生物质材料以包裹投加或直接投加的方式加入水体中,但少有探讨生物质的粒径和投加方式对生物质供碳降氮效果的影响。王新刚等[40]还发现植物在水中腐烂会使次年出水中磷质量浓度增加0.10~0.15 mg·L−1,因此,还需考虑生物质添加对水体磷含量的影响。
本研究将不同粒径的生物质颗粒以无纺布包裹投加和直接投加2种方式对总氮超标的水样进行了处理,考察了在21 d内水体碳、氮、磷含量的变化情况及对总氮的去除效果,以期为利用生物质进行污水与富营养水体的除氮提供参考。
-
WZM球磨机(宜兴市丁蜀浩强机械厂),UV-1800PC紫外可见分光光度计(上海美谱达仪器有限公司),V-1100D可见分光光度计(上海美谱达仪器有限公司),YX-280D压力蒸汽灭菌锅(合肥华泰),FA2004电子天平(上海良平仪器仪表有限公司),干燥器(蜀牛),ZEISS Axioskop40立式显微镜(蔡司公司)。
钼酸铵(AR)、抗坏血酸(AR)、酒石酸锑钾(AR)、浓盐酸(AR)、硫酸锌(AR)、酒石酸钾钠(GR)、苯酚(AR)、氨水(AR)、氢氧化铝(AR)、磷酸(AR)、对氨基苯磺酰胺(AR)、N-(1-萘基) -乙二胺盐酸盐(AR)、浓硫酸(AR)、氢氧化钠(AR)购自成都科隆化学品有限公司,过硫酸钾(AR)购自默克股份两合公司,纳氏试剂购自成都鼎盛时代科技有限公司。
-
稻壳是一种常见的农业固体废弃物,年产量约4 000×104 t。稻壳富含木质素、纤维素、半纤维素、淀粉、还原糖等有机成分,可为微生物提供碳源。相较于木屑、花生壳、甘蔗渣、稻草等农业废弃物,稻壳具有碳源释放能力更稳定、更持久的优势,且稻壳表面粗糙,呈现出网状、针状结构,有利于微生物附着[41]。因此,本研究选用稻壳作为生物质材料。用球磨机将稻壳磨成粉末,分别过20、40、60、80、100、120、140、160目标准筛,从而得到不同粒径的稻壳。
-
无纺布具有吸水、透水能力,且价格低廉、环境友好,可用于包裹生物质或红砖避免泄漏。在生物质释放试验中,取8 cm×8 cm的JS-150A无纺布,在其中心放置1 g稻壳或1 g红砖,对角内折后用橡皮筋捆绑避免散开;在富营养水体脱氮实验中,取15 cm×15 cm的JS-150A无纺布包裹10 g稻壳或红砖。
-
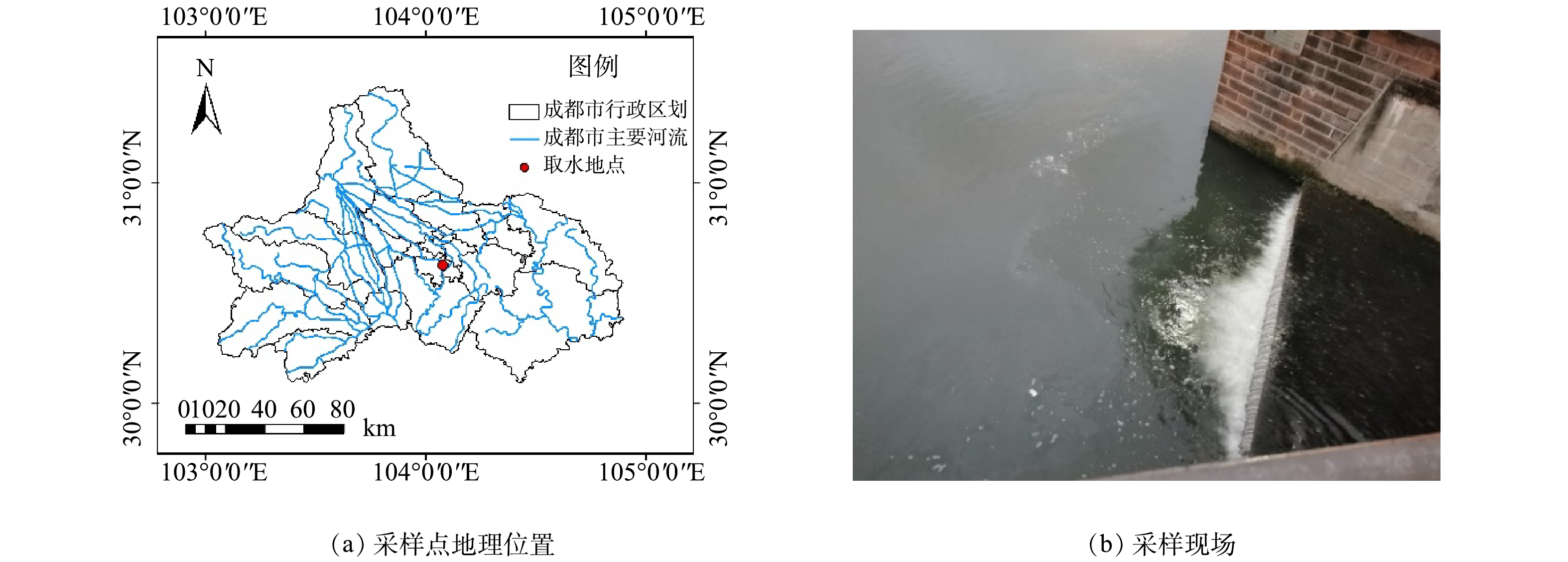
府河是四川省成都市的主要河流之一,自郫都区流入,从彭山县流出,全长115 km,流域面积为2 090 km2,年平均流量可达48 m3·s−1[42]。本研究在府河下游一生活污水排污口处(104.091°E,30.630°N)取得水样,水面有明显油膜(图1),经检测其基本污染物质量浓度情况如表1所示。根据《地表水环境质量标准》(GB 3838-2002),该水样TN质量浓度甚至已超过V类水标准(2.0 mg·L−1),为河水中的主要污染物质。
-
1)生物质释放实验。将稻壳分为20~40、40~80、80~160目3组,其中40~80目组为40~60目与60~80目稻壳按照质量比1:1混合均匀,80~160目组为80~100、100~120、120~140、140~160目稻壳按照质量比1:1:1:1混合均匀。取粒径为20~40、40~80、80~160目的稻壳1 g,分别以直接投加和8 cm×8 cm无纺布包裹的形式投加至1 L超纯水中,搅拌均匀后静置3 d,取上清液检测。以超纯水和(超纯水+无纺布包裹1 g红砖)作为对照。各处理组均设2次重复。
2)富营养水体脱氮实验。将府河水样混合均匀后分为8份10 L水样置于亚克力桶中,分别命名为CK-A0、A1、A2、A3,CK-B0、B1、B2、B3,按照表2添加材料并搅拌均匀。红砖是以黏土、页岩、煤矸石等为原料经高温烧结制成的建筑砖块,稳定性强且比表面积小,有助于无纺布包裹沉入水中,可减小与实验组的差异。各组水样在室外露天条件放置21 d,每隔7 d以虹吸方式取约500 mL水样进行检测并利用显微镜观察水中微藻生长情况。
-
TN、
$ {\text{NH}}_{\text{4}}^{\text{+}}\text{-N} $ 、$ {\text{NO}}_{\text{2}}^{-}\text{-N} $ 、$ {\text{NO}}_{\text{3}}^{-}\text{-N} $ 、TP和CODMn按照水和废水的国家标准方法进行测定[43-44]。有机氮(ON)的浓度用总氮浓度减去无机氮浓度($ \left[\text{ON}\right]\text{=}\left[\text{TN}\right]{-}\left[{\text{NH}}_{\text{4}}^{\text{+}}\text{-N}\right]{-}\left[{\text{NO}}_{\text{2}}^{-}\text{-N}\right]\text{-[}{\text{NO}}_{\text{3}}^{-}\text{-N]} $ [45]) 。 -
1)水样碳、氮、磷含量的计算。在富营养水体脱氮实验中,水样体积受气候影响不断变化,水样中的碳、氮、磷的质量浓度难以直接表现水中碳、氮、磷的变化,采用式(1)计算水样碳、氮、磷的含量。
式中:MX为碳、氮、磷在水中的总质量,mg;
$ {{C}}_{{t}} $ 为t时的CODMn、TP、TN、$ {\text{NH}}_{\text{4}}^{\text{+}}\text{-N}\text{、}{\text{NO}}_{\text{2}}^{-}\text{-N}\text{、} {\text{NO}}_{\text{3}}^{-}\text{-N} $ 、ON质量浓度,mg·L−1;$ {{V}}_{{t}} $ 为t时的水样体积,L;t为取样时间,d。2)实验组水样碳、磷增量的计算。实验中由于生物质的添加,水样碳、磷含量上升,采用式(2)计算实验组相较于对照组的碳、氮、磷质量的增量。
式中:
$ {\Delta }{{M}}_{\text{X}} $ 为实验组水样高于对照组水样碳、磷的质量,mg;$ {{C}}_{\text{0}} $ 为对照组的CODMn、TP质量浓度,mg·L−1。3)统计分析:用SPSS 21对数据进行统计性检验分析。生物质释放实验的多重比较采用Dunnett's T3法,显著性水平为0.10。本研究中所有数据使用Excel 2016进行处理,用Origin 2019绘制图片。
-
以超纯水为空白对照,各项指标均低于标准的检测范围。向超纯水中加入以无纺布包裹的1 g红砖,检测出
$ {\text{NO}}_{\text{3}}^{-} $ 质量浓度仅为0.02~0.03 mg·L−1,CODMn、TP、TN、$ {\text{NH}}_{\text{4}}^{\text{+}}\text{-N} $ 和$ {\text{NO}}_{\text{2}}^{-}\text{-N} $ 均低于标准检测范围,但红砖仍有金属离子溶出的可能。刘海弟等[46]将5 g红砖粉末投入50 mL溶液中,检测发现红砖经纯水浸出的重金属离子 (锰、铅、铬、锌、镍离子)的质量浓度极低,其中锰、铅、铬、锌离子的质量浓度均低于《生活饮用水卫生标准》(GB 5479-2006)规定的浓度限值,而本实验中以10 g红砖投入10 L水溶液中,经稀释后金属离子浓度更低,对水体影响可忽略不计。直接将不同粒径的稻壳颗粒加入超纯水中,释放的CODMn、TP、TN、
$ {\text{NO}}_{\text{3}}^{-} $ 均表现为80~160目>40~80目>20~40目的规律(图2)。这是因为机械粉碎破坏了纤维结构且可增大稻壳的比表面积,从而有利于稻壳的物质释放[47]。邵留等[48]和张雯[49]研究发现,稻壳在3 d内表现出较快的碳、$ {\text{NH}}_{\text{4}}^{\text{+}} $ 和$ {\text{NO}}_{\text{3}}^{-} $ 释放速率,3 d后$ {\text{NH}}_{\text{4}}^{\text{+}} $ 和$ {\text{NO}}_{\text{3}}^{-} $ 释放量减少,且水体碳浓度趋于稳定。因此,认为本研究中在3 d内稻壳已基本完成释放。狄军贞等[50]以5、14、32、60、100目的甘蔗渣进行释碳实验,还原糖和COD释放量随甘蔗渣粒径的减小而先缓慢降低,在32目后迅速升高,与本研究结果相似。小粒径生物质具有更大的表面积,被投加入水中后固-液接触面积更大,此时有利于生物质与水体进行物质交换,进而提高有机物的释放速率[39]。而$ {\text{NH}}_{\text{4}}^{\text{+}} $ 呈现出40~80目组浓度最低的现象,可能是由于不同比表面积下稻壳对$ {\text{NH}}_{\text{4}}^{\text{+}} $ 的吸附和释放平衡不同[51]。另外,仅在直接投加80~160目稻壳颗粒的水中检测到0.013~0.014 mg·L−1的$ {\text{NO}}_{\text{2}}^{-} $ 释放。将稻壳颗粒以无纺布包裹后加入水中,仅对80~160目稻壳的CODMn和$ {\text{NH}}_{\text{4}}^{\text{+}} $ 释放产生了显著影响(P<0.1)。稻壳被包裹后运动范围受限,不利于水在颗粒间的流动,水与稻壳的大分子物质交换受阻,且由于80~160目稻壳颗粒间空隙更小,这种影响更显著。稻壳释放的CODMn/TN表现为80~160目>40~80目>20~40目(P>0.1)、直接投加>无纺布包裹投加(P<0.1)的规律(图2)。直接投加时稻壳释放的CODMn/TN为12.4~19.7,用无纺布包裹投加后稻壳释放的CODMn/TN为9.8~12.7,与稻草、梭鱼草、绿狐尾藻释放的COD/TN相近[41],且高于常规认为反硝化所需的C/N[20-21],因此,认为在直接投加和无纺布包裹投加的方式下,不同粒径的稻壳均能满足提供碳源、提高水体C/N促进反硝化的要求。稻壳释放的CODMn/TP为27.9~37.5,且未表现出受稻壳粒径和投加方式的显著影响。
-
在实验过程中,对照组碳含量(MCOD)不断波动。由于微生物代谢消耗,CK-A0与CK-B0在第7天有机物含量减少至20.8~21.7 mg,至14 d由于水中浮游藻类浓度较高而表现出较高的有机物含量(54.3~57.8 mg),至21 d有机物含量又降至初始水平附近(图3)。添加稻壳后的水样始终表现出较高的碳增量(
${\Delta }{{M}}_{\text{COD}}$ )。任玉锐等[29]用2 g橘树叶、玉米芯进行为期19 d的反硝化实验,但在15 d后碳源释放量开始减少,脱氮效果下降。本研究中稻壳表现出持久的释碳能力,实验组${\Delta }{{M}}_{\text{COD}}$ 持续升高且在第21天达到最高值(41.1~107.3 mg)。然而实验组${\Delta }{{M}}_{\text{COD}}$ 始终低于稻壳直接释放的碳含量,这表明有微生物利用和消耗了水中的碳源。粒径对水样
$ {\Delta }{\text{M}}_{\text{COD}} $ 有明显影响,表现为80~160目>40~80目>20~40目。其中直接投加80~160目时水样碳增量高达90.4~107.3 mg,远高于其他实验组(36.1~65.6 mg)(图3)。因此,添加小粒径颗粒可达到快速改变水体营养元素化学计量比的效果,但这种快速调节的优势也伴随着碳源不能持久供给的问题[52]。投加方式对水样
$ {\Delta }{\text{M}}_{\text{COD}} $ 也有明显影响,且$ {\Delta }{\text{M}}_{\text{COD}} $ 表现为直接投加组>无纺布包裹投加组,且两者间差异大于超纯水中稻壳释放的CODMn差异,应为微生物分解稻壳的影响。以无纺布包裹稻壳,稻壳间空隙减小,不利于微生物附着形成生物膜[53],微生物对稻壳的分解利用减慢。因此用无纺布包裹稻壳不仅可以减缓稻壳与水的物质交换,还可以减缓微生物对稻壳的分解作用,达到长久供碳的效果。目前常用于调节生物质的碳供给的方法有以下3类:采用酸处理、碱处理、双氧水处理、紫外线照射等物理、化学方法对生物质进行预处理,其主要通过破坏木质素保护层和纤维素的晶格结构改变生物质释放营养物质的速率、种类[54-56];将生物质在高温条件下经厌氧处理碳化成为一种富碳物质,不仅可减少生物质中可利用生物碳,避免了出水高色度和高有机物浓度的问题,且改变了生物质孔径、比表面积、表面活性基团等物理化学性质,有利于污染物质的吸附和微生物的附着[57-59];将生物质与聚乙烯、沸石粉、膨润土、高岭土等物质混合,制成具有一定硬度和抗压强度的填料载体,减缓营养物质的释放速率[55, 60-61]。但是,这些处理方式复杂、技术难度大、处理成本高,不利于工业大规模生产和利用。本研究通过改变生物质粒径和投加方式的简单物理处理方法实现了生物质碳供给速率的调节,具有应用优势。
-
由图4可见,实验过程中,对照组水样的磷含量(MTP)持续下降。对照组的MTP由于微生物消耗而持续降低,降低速度在第7~14天最快,由0.99~1.03 mg下降至0.30~0.34 mg,期间浮游藻类浓度为实验期间最高水平。此后对照组磷的消耗速率和浮游藻类浓度都下降至较低水平,至第21天MTP为0.16~0.25 mg。实验组水样的MTP在第7天达到最高(2.13~3.35 mg),此后持续下降。
实验组始终具有磷增量(
$ {\Delta }{\text{M}}_{\text{TP}} $ ),且各实验组的$ {\Delta }{\text{M}}_{\text{TP}} $ 均在第7天达到最高(1.14~2.32 mg),此后逐渐减小。但本实验中$ {\Delta }{\text{M}}_{\text{TP}} $ 始终低于稻壳直接释放的磷含量,且$ {\Delta }{\text{M}}_{\text{TP}} $ 持续减小证明实验组的磷消耗速率快于对照组,这可能是因为磷酸盐在细胞表面被吸附并逐渐被同化,最终随着微生物沉降,且微藻光合作用使水体pH上升有利于磷酸盐的沉降[62]。本实验中粒径对水样$ {\Delta }{\text{M}}_{\text{T}\text{P}} $ 有明显影响,$ {\Delta }{\text{M}}_{\text{T}\text{P}} $ 大小顺序为80~160目组>40~80目组>20~40目组,而投加方式对$ {\Delta }{\text{M}}_{\text{T}\text{P}} $ 的影响不明显。 -
图5为以包裹投加和直接投加的方式添加不同粒径的稻壳后,水样中各形态氮含量变化情况。2对照组TN含量在0~7 d较为稳定,在7~14 d水中MTN由29.4~39.1 mg快速下降至14.2~14.6 mg,14~21 d水中各形态氮含量均保持稳定,其中
$ {\text{NO}}_{\text{2}}^{-} $ 含量始终维持在较高水平(>0.9 mg)且持续上升,可能为低C/N比下硝化-反硝化作用受阻导致$ {\text{NO}}_{\text{2}}^{-} $ 积累,ON则被转化为其他形态氮,使得MON由11.5~11.7 mg降至9.5~10.6 mg(图5)。实验组水样中氮的含量和存在形态与对照组不同。7 d后MTN略有上升且MON(14.1~17.7 mg)均高于对照组(11.4~11.6 mg),可能为稻壳释放
$ {\text{NH}}_{\text{4}}^{\text{+}} $ 和蛋白质、氨基酸等含氮有机物所致,A3组中由于稻壳释放$ {\text{NO}}_{\text{2}}^{-} $ 和缺氧环境中硝化受阻,导致$ {\text{NO}}_{\text{2}}^{-} $ 升高至2.7 mg,远大于其他实验组(1.0~1.5 mg)。丁绍兰等[63]和朱辉翔等[64]以核桃壳、花生壳、稻草等生物质作为反硝化碳源时,实验前期也出现$ {\text{NO}}_{\text{2}}^{-} $ 积累现象。7~21 d,实验组MTN为10.6~7.5 mg·L−1,低于对照组(13.2~14.6 mg·L−1);$ {\text{NO}}_{\text{2}}^{-} $ 含量均小于0.15 mg,远低于对照组;MON迅速下降至6.7~11.7 mg,消耗速率较对照组更快。第14~21天,实验组各形态氮含量维持稳定。多数研究以生物质作为反硝化碳源加入人工配置的污水中,并接种活性污泥,达到了良好的氮去除效果。如朱辉翔等[64]以稻草、绿狐尾藻处理用KNO3、KH2PO4配置的污水并接种活性污泥,于第5天
$ {\text{NO}}_{\text{3}}^{-} $ 去除率达到90%以上;丁绍兰等[63]以壳类生物质处理用KNO3、K2HPO4配置的污水并接种活性污泥,水中$ {\text{NO}}_{\text{3}}^{-} $ 去除率在2~12 d均维持在70%~85%。本研究从自然水中取样,水中氮主要以ON和$ {\text{NH}}_{\text{4}}^{\text{+}} $ 的形式存在,加入稻壳后水中TN和ON的去除率至21 d分别达到了71.1%~75.1%和60.1%~66.2%,高于对照组的53.6%~56.1%、43.9%~49.6%,而$ {\text{NH}}_{\text{4}}^{\text{+}} $ 与对照组$ {\text{NH}}_{\text{4}}^{\text{+}} $ 均接近于0。其次,本研究利用水体中自然存在的微生物,而未接种活性污泥,因此,在实验前期微生物需要适应新的水体环境,这可能也是至第7天实验组仍未表现出较好脱氮效果的原因。MULLER[65]以PHA(聚-β-羟基-丁酸)颗粒作为反硝化碳源和微生物附着基质利用,当PHA颗粒的表面积增加1倍时,反硝化速率几乎也增加1倍。在本研究中,以直接投加或以无纺布包裹投加不同粒径的稻壳颗粒时,未观察到总氮去除效果与生物质粒径(表面积)或投加方式的显著相关性,但在实验前期水中氮的存在形态有所差异,如第7天直接添加40~80目稻壳颗粒的$ {\text{NO}}_{\text{3}}^{-} $ 含量、直接添加80~160目稻壳颗粒的$ {\text{NO}}_{\text{2}}^{-} $ 含量均明显高于其他实验组。这可能是不同C/N下微生物的适应性不同,随着时间延长,氮存在形态的差异也逐渐减小。 -
大量污废水存在营养元素比例失调的问题,水中微生物需要通过减少生长繁殖或加剧能量消耗来调节体内营养平衡[66],这使水体自净过程受到阻碍。STUTTER[67]提出,当水体的C/N为2~11,C/P为47~994时,水中营养元素比例较为平衡,异养微生物能适应水体环境并有效利用氮、磷。本研究中选取水样的初始CODMn:TN:TP比例为24.2:25.9:1,该比例已脱离微生物代谢的舒适区,P和N饱和且与C循环脱钩,导致氮去除困难。生物质包含纤维素、果糖、木质素等含C有机物质,蛋白质、生物碱等含N物质和核酸、磷脂等含P物质,不同种类生物质的元素和物质比例各有不同。加入稻壳后,葡萄糖、蛋白质、脂肪酸等快速进入水中并被微生物分解利用,使水中C、N、P浓度迅速上升、比例改变。水样CODMn/TN由0.935上升至8.5~16.5,远高于对照组的0.5~4(图6),但略低于稻壳直接释放的CODMn/TN(8.5~19.7)(图2)。这是因为水中有微生物利用碳源进行分解。水样CODMn/TP不断升高至170以上(图6),远高于稻壳在超纯水中直接释放的CODMn/TP(27.9~37.5)(图2)。这是因为稻壳的添加补充了水中碳源,且水中磷大量被微生物消耗或沉降至水底。此时水体环境更利于微生物生长并利用氮、磷。稻壳的纤维素、半纤维素、木质素这类结构物质分解速度则相对缓慢,尤其木质素对生物分解有很强的抵抗力[68],使生物质表现出更长效的供碳效果,因此,水样CODMn/TN和CODMn/TP在21 d内仍维持在较高水平。
本实验中生物质的粒径和投加方式影响水体碳、氮、磷含量变化而导致CODMn/TN、CODMn/TP改变。至第21天,水体的CODMn/TN呈现出80~160目>40~80目>20~40目、直接投加组>无纺布包裹投加组的规律,而CODMn/TP则呈现出20~40目>40~80目>80~160目、直接投加组>无纺布包裹投加组的规律。
-
水中浮游植物(尤其是藻类)的数量迅速增加是水体富营养化的显著特征之一,浮游植物结构和物种多样性的变化可以反映水质的变化[13,69]。藻类是促进水体氮去除的重要生物,一般以Redfield比率(C:N:P=106:16:1)[70]作为藻类生长的最佳值。从化学计量比的角度分析,本研究中对照组N含量偏高不利于藻类生长,实验组的C含量偏高,可能因为稻壳添加量高于适宜值。
资源比率理论[69](resource-ratio theory)认为随着水体中不同元素比例的变化,具有不同最佳养分比例的藻类相互竞争而导致群落结构发生变化,如HILLEBRAND等[71]研究表明,硅藻的最佳N:P为14.9,而对于甲藻和蓝藻则分别为15.1和25.8。本研究在800倍显微镜下观察发现,在所有水样中均有黄藻门的黄丝藻、硅藻门的针杆藻和绿藻门的栅藻等,但在CODMn/TN、CODMn/TP更高的实验组水样中观察到数量更多的针杆藻和栅藻等,且还观察到硅藻门的舟形藻、绿藻门的红球藻等,这说明浮游植物种类更丰富(图7)。于潘等[72]使用光学显微镜对微藻样品进行了鉴定和计数。但本研究中藻类浓度较低,难以直接通过显微镜观察计数,且水样有限,为了减少对实验结果的影响,无法取样浓缩用以藻类计数。ISHIDA等[73]研究发现硅藻与反硝化间存在正相关关系,推测硅藻与反硝化菌间存在协同关系。由于藻类不仅可以直接吸收利用
$ {\text{NH}}_{\text{4}}^{\text{+}}\text{、}{\text{NO}}_{\text{3}}^{-} $ ,且在日间可为水中硝化、氨氧化菌群提供氧气促进代谢,在夜间消耗氧气为反硝化提供低氧环境[65,73],实验组均表现出优于对照组的TN、$ {\text{NO}}_{\text{2}}^{-} $ 、ON去除率。 -
1)稻壳的粒径决定稻壳的比表面积,从而影响稻壳与水体的物质交换速率,导致水体的碳、氮、磷含量及比例存在差异。当粒径为80~160目时,水中固-液面积大,有利于稻壳与水体的物质交换,释放的碳、氮、磷含量最高。但将稻壳应用于富营养水体脱氮时,粒径仅在短期内使水体氮存在形态表现出差异,而从长期来看对水体中氮的去除效果无影响。水中碳、磷含量变化使CODMn/TN呈现出随粒径减小而升高的趋势,CODMn/TP呈现出随粒径减小而下降的趋势。
2)稻壳的投加方式决定了稻壳颗粒间的孔隙度,影响稻壳与水体的碳交换速率和微生物附着生长,导致水体的碳含量及碳氮磷比例存在差异。直接投加80~160目稻壳时,稻壳释放的碳含量高于用无纺布包裹投加相同粒径的稻壳,而氮、磷含量无显著差异,将其应用于富营养水体氮去除时也表现出相同的结果。此时水中CODMn/TN、CODMn/TP仅受碳含量影响,直接投加生物质时水体CODMn/TN、CODMn/TP均高于用无纺布包裹投加。
生物质利用方式对富营养水体中碳氮磷含量及其比例的影响
Effects of biomass utilization on carbon, nitrogen and phosphorus contents and their proportion in eutrophic water
-
摘要: 生物质常被添加至富营养水体中以调节碳氮比(C/N),促进脱氮,但不清楚生物质的粒径和投加方式对富营养水体中营养物质浓度和比例的影响。通过机械粉碎制得20~40、40~80、80~160目稻壳颗粒,并选取直接投加和无纺布包裹投加2种投加形式,以不添加稻壳或无纺布包裹砖块作为对照组。以超纯水为水样研究不同粒径和投加方式下稻壳的3 d碳氮磷释放特性;以成都市府河下游河段水样为研究对象,考察了不同粒径和投加方式下稻壳的21 d静态脱氮特性。结果表明,在超纯水中,稻壳释放出CODMn(5.5~16.5 mg·L−1)、TP(0.18~0.45 mg·L−1)、TN(0.39~0.95 mg·L−1)、CODMn/TN(8.5~19.7)、CODMn/TP(27.9~37.5),前四者均呈现出80~160目>40~80目>20~40目的规律,仅CODMn和CODMn/TN呈现直接投加>无纺布包裹投加的规律。在静态脱氮实验中,21 d后,添加了稻壳的水样CODMn含量由28.1 mg上升至71.7~143.0 mg,TP含量由1.2 mg下降至0.2~0.5 mg,CODMn/TN由0.9上升至8.5~16.5,CODMn/TP由24.2上升至170以上,这4个参数的变化情况与稻壳的碳氮磷释放规律相同;TN含量由30.04 mg下降至7.64~8.68 mg,低于对照组(13.2~13.9 mg),但粒径和投加方式仅对实验初期水中氮的存在形态有影响。粒径决定稻壳的比表面积,投加方式决定稻壳的间隙,并影响微生物的附着效果,进而影响稻壳的碳氮磷释放效果。在实际应用时可根据情况,采用不同的生物质粒径和投加方式,以调节水中营养物质浓度和比例。Abstract: Biomass is often added to eutrophic water to adjust the ratio of carbon to nitrogen ratio (C/N) and promote denitrification. However, the effects of particle size and dosing mode on the nutrient concentrations and their proportion in the eutrophic water are not clear. In this study, rice husk with the particle size of 20~40, 40~80 and 80~160 mesh was prepared by mechanical grinding, and two modes such as directly and non-woven fabrics-wrapped dosing were used to conduct the experiments with control groups of adding nothing or bricks wrapped with non-woven fabric. The carbon, nitrogen and phosphorus release characteristics of rice husk under the conditions of different grain sizes and dosing modes in 3 days were studied by using ultra-pure water as water samples, while the static nitrogen removal characteristics of rice husk with different grain sizes and dosing modes in 21 days were studied by using water samples from the lower reaches of Fuhe River in Chengdu. The results showed that in the ultra-pure water, rice husk released CODMn of 5.5~16.5 mg·L−1, TP of 0.18~0.45 mg·L−1 and TN of 0.39~0.95 mg·L−1, CODMn/TN ratio of 8.5~19.7, CODMn/TP ratio of 27.9~37.5, and the first four indices showed the order: 80~160 mesh >40~80 mesh >20~40 mesh, while only CODMn and CODMn/TN showed the order: direct addition > non-woven fabric-wrapped addition. In the static nitrogen removal experiments with rice husks-dosed water samples after 21 days, CODMn increased from 28.1 mg to 71.7~143.0 mg, TP decreased from 1.2 mg to 0.2~0.5 mg, CODMn/TN increased from 0.9 to 8.5~16.5, CODMn/TP increased from 24.2 to more than 170, which showed the same trends as above rice husk release experiments in ultra-pure water; TN decreased from 30.04 mg to 7.64~8.68 mg, which was lower than that of the control group (13.2~13.9 mg), but the particle size and dosing modes of rice husks only affected N species at the beginning of the experiment. The particle size of rice husks determined its specific surface, the dosing modes determined the gap between rice husks, they affected the attachment effect of microorganisms, and then the release of carbon, nitrogen and phosphorus. In actual application, the various combination of the biomass particle size and non-woven fabric-wrapping can be used to adjust the concentration and proportion of nutrients in water.
-
Key words:
- eutrophic water /
- biomass /
- carbon /
- nitrogen /
- phosphorus
-

-
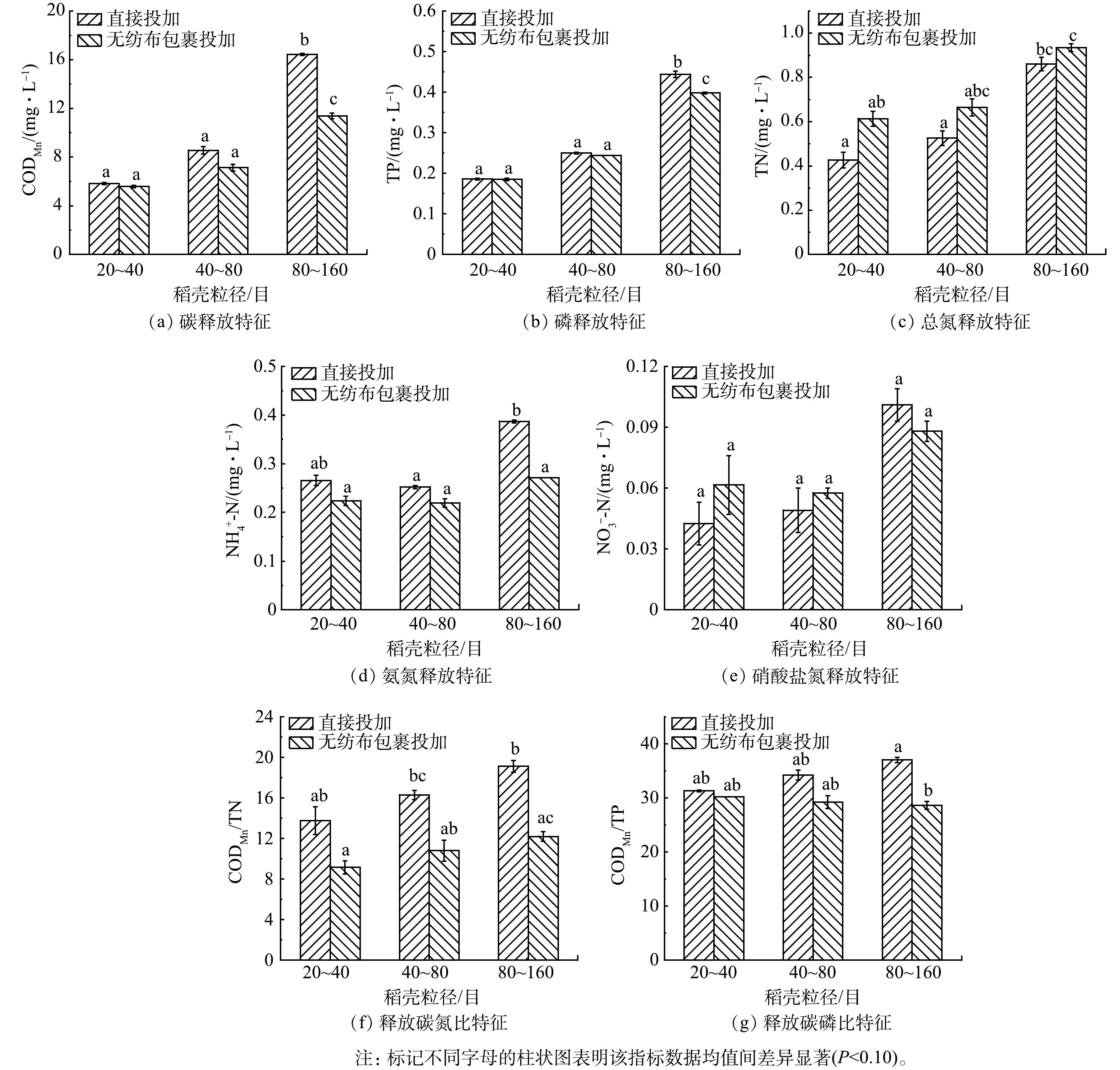
图 2 直接投加和包裹添加不同粒径稻壳时稻壳的碳氮磷释放特征
Figure 2. The release of carbon, phosphorate and nitrogen after directly and wrapped dosing of rice husk with different particle sizes
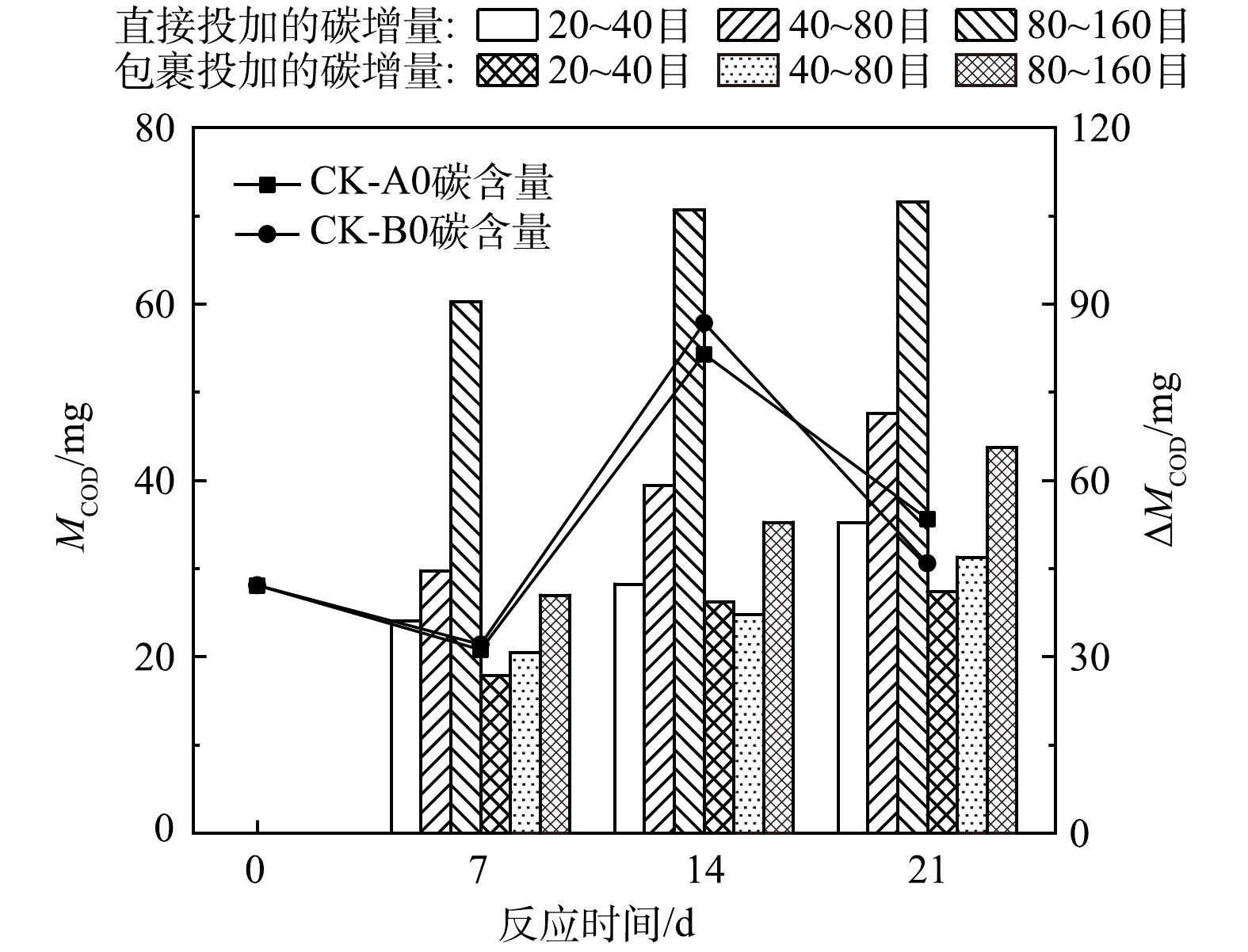
图 3 直接投加和包裹添加不同粒径稻壳后水体碳含量变化特征
Figure 3. Variation characteristics of carbon quality in water after directly and wrapped dosing of rice husk with different particle sizes
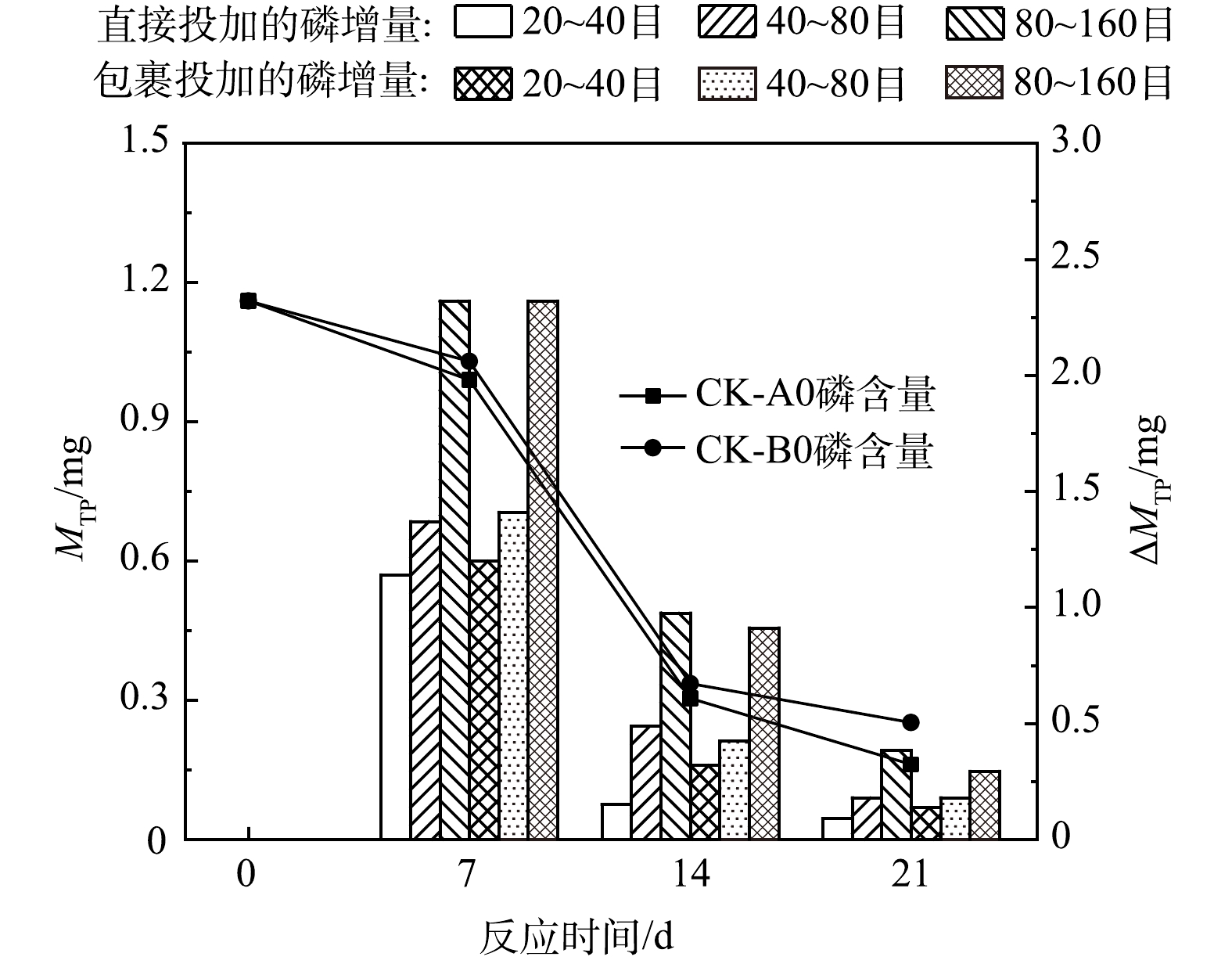
图 4 直接投加和包裹添加不同粒径稻壳后水体磷含量变化特征
Figure 4. Variation characteristics of phosphorus quality in water after directly and wrapped dosing of rice husk with different particle sizes
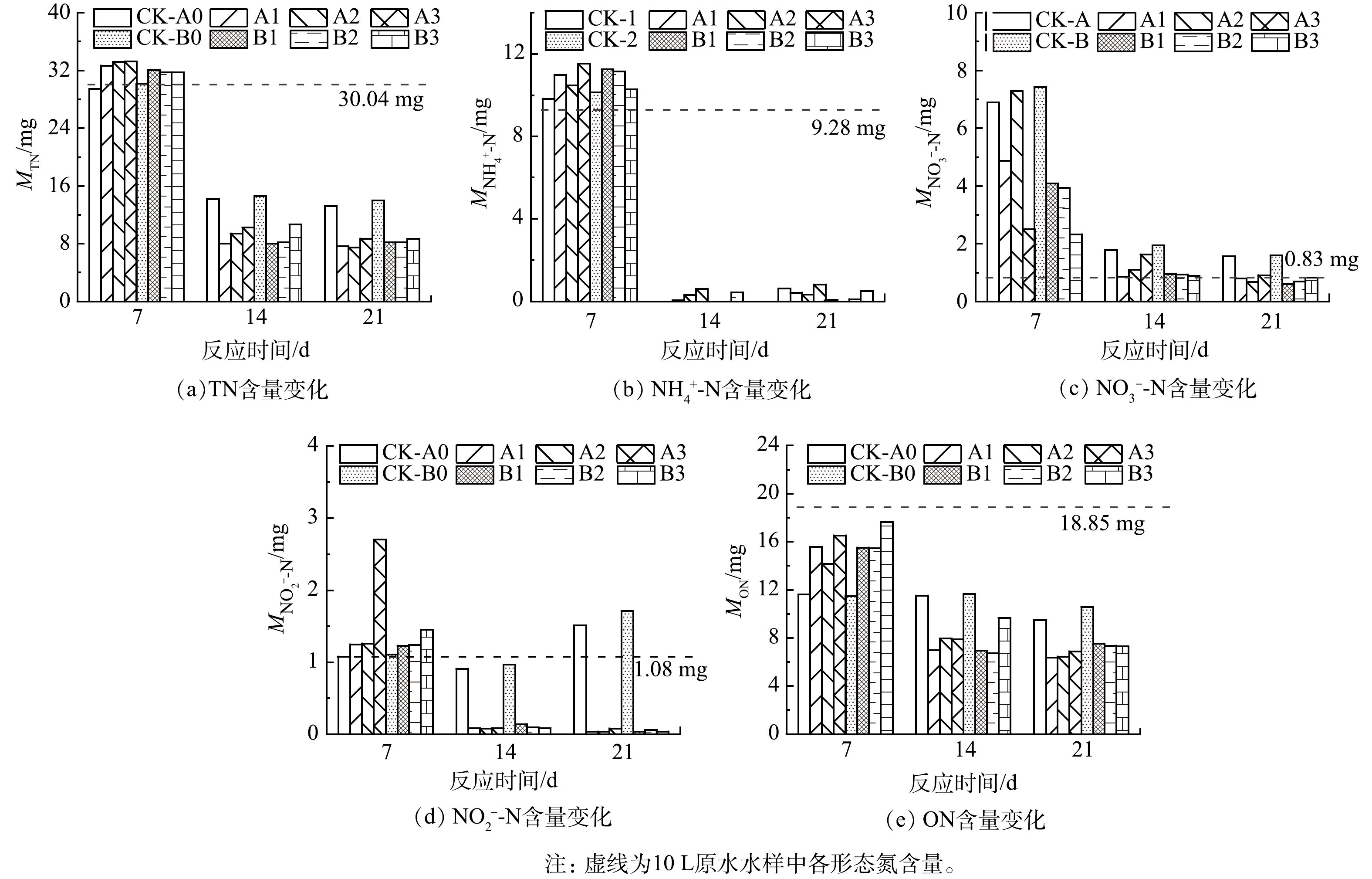
图 5 直接投加和包裹添加不同粒径稻壳后水体中氮的去除特征
Figure 5. Variation characteristics of nitrogen quality in water after directly and wrapped dosing of rice husk with different particle sizes
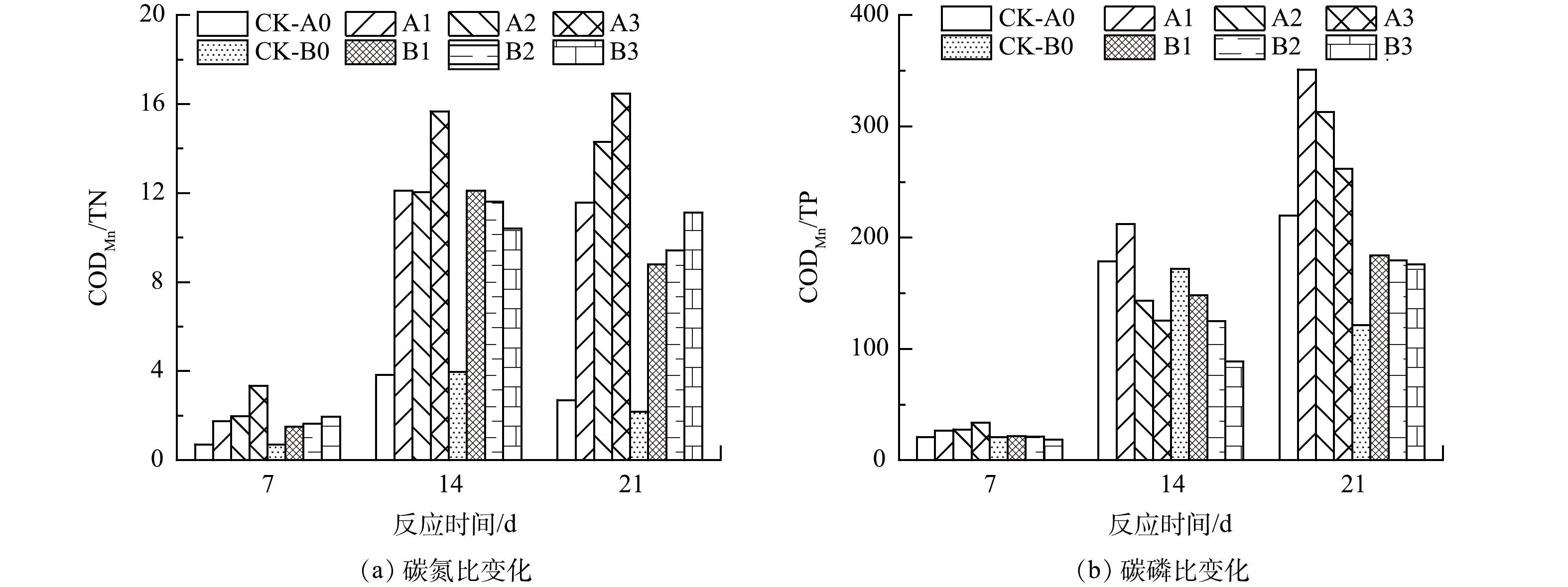
图 6 直接投加和包裹添加不同粒径稻壳时水体的碳氮磷比例变化特征
Figure 6. Variation characteristics of carbon, nitrogen and phosphorus ratio in water after directly and wrapped dosing of rice husk with different particle sizes

图 7 直接投加和包裹添加不同粒径稻壳时水中微藻生长情况
Figure 7. Microalgae growth in water after directly and wrapped dosing of rice husk with different particle sizes
表 1 水样基础数据
Table 1. Basic data of water samples
mg·L−1 样品 TP TN NH4+-N NO2−-N NO3−-N CODMn GB 3838-2002 III类水标准 0.2 1.0 1.0 — — 4 原水水样 0.17 3.00 0.93 0.108 0.08 2.8 注:“—”表示标准中无该项指标。  下载: 导出CSV
下载: 导出CSV
表 2 实验处理
Table 2. Experimental treatment
水样 水样/L 稻壳/g 红砖/g 是否使用
无纺布包裹20~40目 40~80目 80~160目 CK-A0 10 — — — — 否 A1 10 10 — — — 否 A2 10 — 10 — — 否 A3 10 — — 10 — 否 CK-B0 10 — — — 10 是 B1 10 10 — — — 是 B2 10 — 10 — — 是 B3 10 — — 10 — 是 注:“—”表示水样中未添加该材料。
下载: 导出CSV
-
[1] MOAL M L, GASCUEL-ODOUX C, MéNESGUEN A, et al. Eutrophication: A new wine in an old bottle?[J]. Science of the Total Environment, 2019, 651(Pt 1): 1-11. [2] LE C, ZHA Y, LI Y, et al. Eutrophication of lake waters in China: Cost, causes, and control[J]. Environmental Management, 2010, 45(4): 662-668. doi: 10.1007/s00267-010-9440-3 [3] 丁雄祺, 谢媚, 陈偿, 等. 一株高效氨氮及亚硝态氮去除功能菌株的分离鉴定及在生物絮团对虾养殖中的应用[J]. 中国水产科学, 2019, 26(5): 959-970. [4] 孔繁翔, 高光. 大型浅水富营养化湖泊中蓝藻水华形成机理的思考[J]. 生态学报, 2005, 25(3): 589-595. doi: 10.3321/j.issn:1000-0933.2005.03.028 [5] NILOUFAR G, R. P K. Critical review of effluent dissolved organic nitrogen removal by soil/aquifer-based treatment systems[J]. Chemosphere, 2021, 269: 1-11. [6] WANG H, WANG T, YANG S, et al. Nitrogen removal in oligotrophic reservoir water by a mixed aerobic denitrifying consortium: Influencing factors and immobilization effects[J]. International Journal of Environmental Research and Public Health, 2019, 16(4): 583. doi: 10.3390/ijerph16040583 [7] 张文艺, 张采芹, 占明飞, 等. 滤食性底栖动物-菌藻复合生态系统对富营养水体净化的特性[J]. 湖北农业科学, 2013, 52(20): 4926-4931. [8] 朱小龙, 谷娇, 靳辉, 等. 太湖河蚬(Corbicula fluminea)对富营养水体水质的改善作用?[J]. 湖泊科学, 2015, 27(3): 486-492. doi: 10.18307/2015.0316 [9] 江君, 李欣, 徐飞, 等. 3个荷花品种对富营养水体和底泥中氮、磷去除能力比较研究[J]. 江苏农业科学, 2018, 46(8): 296-299. [10] 徐恒戬, 权召, 周永顺. 富营养水体修复植物种质筛选的研究[J]. 种子, 2018, 37(5): 67-69. [11] 张萌, 李雄清, 邹新, 等. 典型沉水植物修复富营养水体的最优种植密度[J]. 湖北农业科学, 2016, 55(20): 5218-5224. [12] 梁玉婷, 杨星宇, 杨兰芳, 等. 湿地植物生长对去除富营养化水体总氮和硝氮的影响[J]. 湖北大学学报(自然科学版), 2021, 43(6): 644-652. [13] LIN S S, SHEN S L, ZHOU A, et al. Assessment andmanagement of lake eutrophication: A case study in Lake Erhai, China[J]. Science of the Total Environment, 2021, 751: 2021, 751: 141618 [14] XINGHUI X, SIBO Z, SILING L, et al. The cycle of nitrogen in river systems: sources, transformation, and flux[J]. Environmental science Processes & impacts, 2018, 20(6): 863-891. [15] KUYPERS M M M, MARCHANT H K, KARTAL B. The microbial nitrogen-cycling network[J]. Nature Reviews Microbiology, 2018, 16(5): 263-276. doi: 10.1038/nrmicro.2018.9 [16] GAO F, YANG H-L, LI C, et al. Effect of organic carbon to nitrogen ratio in wastewater on growth, nutrient uptake and lipid accumulation of a mixotrophic microalgae Chlorella sp[J]. Bioresource Technology, 2019, 282: 118-124. doi: 10.1016/j.biortech.2019.03.011 [17] SAEED T, GUANGZHI S. A review on nitrogen and organics removal mechanisms in subsurface flow constructed wetlands: dependency on environmental parameters, operating conditions and supporting media[J]. Journal of Environmental Management, 2012, 112: 429-448. doi: 10.1016/j.jenvman.2012.08.011 [18] 廖晓数, 贺锋, 徐栋, 等. 低C/N对湿地中硝化反硝化作用的影响[J]. 中国环境科学, 2008, 28(7): 603-607. doi: 10.3321/j.issn:1000-6923.2008.07.006 [19] 吴代顺, 杨昕怡, 于雪, 等. 碳氮比对硝化过程微生物代谢及功能基因的影响[J]. 中国给水排水, 2021, 37(7): 20-26. [20] BONASSA G, BOLSAN A C, HOLLAS C E, et al. Organic carbon bioavailability: Is it a good driver to choose the best biological nitrogen removal process?[J]. Science of the Total Environment, 2021, 786: 147390. doi: 10.1016/j.scitotenv.2021.147390 [21] CHIU Y C, CHUNG M S. Determination of optimal COD/nitrate ratio for biological denitrification[J]. International Biodeterioration & Biodegradation, 2003, 51(1): 43-49. [22] CAO S B, DU R, PENG Y Z, et al. Novel two stage partial denitrification (PD)-Anammox process for tertiary nitrogen removal from low carbon/nitrogen (C/N) municipal sewage[J]. Chemical Engineering Journal, 2019, 362: 107-115. doi: 10.1016/j.cej.2018.12.160 [23] FU X R, HOU R R, YANG P, et al. Application of external carbon source in heterotrophic denitrification of domestic sewage: A review[J]. Science of the Total Environment, 2022, 817: 153061. doi: 10.1016/j.scitotenv.2022.153061 [24] 王建华, 陈永志, 彭永臻. 低碳氮比实际生活污水A2O-BAF工艺低温脱氮除磷[J]. 中国环境科学, 2010, 30(9): 1195-1200. [25] 胡曼利. 外加植物碳源和稀碱加热预处理强化潜流人工湿地对低C/N污水的处理效果[D]. 重庆: 西南大学, 2022. [26] 彭永臻, 王鸣岐, 彭轶, 等. 四种碳源条件下城市污水处理厂尾水深度脱氮的性能与微生物种群结构[J]. 北京工业大学学报, 2021, 47(10): 1158-1166. [27] CHIU Y C, LEE L L, CHANG C N, et al. Control of carbon and ammonium ratio for simultaneous nitrification and denitrification in a sequencing batch bioreactor[J]. International Biodeterioration & Biodegradation, 2007, 59(1): 1-7. [28] 凌宇, 闫国凯, 王海燕, 等. 6种农业废弃物初期碳源及溶解性有机物释放机制[J]. 环境科学, 2021, 42(5): 2422-2431. [29] 任玉锐, 郭照冰. 植物作为反硝化碳源[J]. 环境工程学报, 2015, 9(5): 2247-2252. doi: 10.12030/j.cjee.20150536 [30] 余晖, 张学青, 张曦, 等. 黄河水体颗粒物对硝化过程的影响研究[J]. 环境科学学报, 2004, 24(4): 601-606. doi: 10.3321/j.issn:0253-2468.2004.04.007 [31] 张学青, 夏星辉, 杨志峰. 水体颗粒物对有机氮转化的影响[J]. 环境科学, 2007, 28(9): 1954-1959. doi: 10.3321/j.issn:0250-3301.2007.09.010 [32] LYNN T J, YEH D H, ERGAS S J. Performance of denitrifying stormwater biofilters under intermittent conditions[J]. Environmental Engineering Science, 2015, 32(9): 796-805. doi: 10.1089/ees.2015.0135 [33] BOLEY A, MULLER W R, HAIDER G. Biodegradable polymers as solid substrate and biofilm carrier for denitrification in recirculated aquaculture systems[J]. Aquacultural Engineering:An International Journal, 2000(1/2): 22. [34] 陈志华, 周键, 王三反. 固相反硝化在水污染治理中的研究进展[J]. 化工进展, 2021, 40(z1): 366-374. [35] 雷柯柯, 马甡, 单洪伟, 等. 人工悬浮生物絮团在凡纳滨对虾养殖系统中的初步应用[J]. 水产科技情报, 2019, 46(1): 37-43. [36] 罗佳, 韩士群, 罗海荣, 等. 外加碳源对富营养化水体生物脱氮效果及细菌群落结构的影响[J]. 江苏农业学报, 2012, 28(6): 1312-1317. [37] 王玥, 秦帆, 唐燕华, 等. 农业废弃物作为反硝化脱氮外加碳源的研究[J]. 林业工程学报, 2019, 4(5): 146-151. [38] 王琦, 石雷, 杨小丽, 等. 废弃生物质强化生态袋脱氮除磷的效果[J]. 东南大学学报(自然科学版), 2021, 51(1): 138-144. doi: 10.3969/j.issn.1001-0505.2021.01.019 [39] 杨正健, 魏辰宇, 刘德富, 等. 开放水体脱氮过程及其影响因素研究进展[J]. 水利学报, 2021, 52(2): 194-202. [40] 王新刚, 吕锡武, 张圣菊. 水生植物分解过程中生物质及氮磷释放规律研究[J]. 安全与环境学报, 2011, 11(5): 82-85. doi: 10.3969/j.issn.1009-6094.2011.05.018 [41] 邵留, 徐祖信, 金伟, 等. 农业废物反硝化固体碳源的优选[J]. 中国环境科学, 2011, 31(5): 748-754. [42] 成都市地方志编纂委员会编纂. 成都市志 水利志[M]. 成都: 四川辞书出版社, 2001. [43] 林浩澎, 孙慧明, 罗娉婷, 等. 一株耐碱变形假单胞菌ZY-3的鉴定及其脱氮特性[J]. 微生物学通报, 2022, 49(10): 4066-4079. [44] 张怡, 刘本洪, 刘蕾, 等. 硬质河岸和水体富营养化河道的综合治理技术: 以柴桑河为例[J]. 环境工程学报, 2021, 15(12): 3875-3882. doi: 10.12030/j.cjee.202109012 [45] 丁润楠, 姚晓龙, 傅大放, 等. 中国东部湖泊有机氮浓度时空特征及影响因素[J]. 环境科学与技术, 2021, 44(6): 35-42. [46] 刘海弟, 孙鲁强, 李伟曼, 等. 红砖烧成对污泥原料中重金属离子固化的研究[J]. 砖瓦, 2021(8): 21-22. doi: 10.3969/j.issn.1001-6945.2021.08.007 [47] 岳建芝, 李刚, 张全国. 促进木质纤维素类生物质酶解的预处理技术综述[J]. 江苏农业科学, 2011, 39(3): 340-343. doi: 10.3969/j.issn.1002-1302.2011.03.134 [48] 邵留, 徐祖信, 王晟, 等. 新型反硝化固体碳源释碳性能研究[J]. 环境科学, 2011, 32(8): 2323-2327. [49] 张雯. 以农业废弃物为基料的地下水反硝化缓释碳源材料研究及应用[D]. 南京: 南京大学, 2017. [50] 狄军贞, 李拓达, 赵微. 甘蔗渣碳源释放规律及其硫酸盐还原菌利用性试验[J]. 农业环境科学学报, 2019, 38(5): 1151-1157. doi: 10.11654/jaes.2018-0994 [51] 阳春, 郑向勇, 严立, 等. 几种土壤改良材料磷氨氮吸附和硝化作用特性的研究[J]. 农业环境科学学报, 2008(5): 2013-2017. doi: 10.3321/j.issn:1672-2043.2008.05.058 [52] SALILING W J B, WESTERMAN P W, LOSORDO T M. Wood chips and wheat straw as alternative biofilter media for denitrification reactors treating aquaculture and other wastewaters with high nitrate concentrations[J]. Aquacultural Engineering, 2007, 37(3): 222-233. doi: 10.1016/j.aquaeng.2007.06.003 [53] 田冬, 高明, 王侃. 不同粒径生物质灰渣填料净化生活污水的试验研究[J]. 水土保持学报, 2015, 29(4): 218-222. [54] 赵文莉, 郝瑞霞, 李斌, 等. 预处理方法对玉米芯作为反硝化固体碳源的影响[J]. 环境科学, 2014, 35(3): 987-994. [55] 杨平, 刘青松, 石广辉, 等. 稻壳作为缓释碳源及载体的改性研究[J]. 生态科学, 2019, 38(2): 112-118. [56] 李斌. 固体生物质碳源复合滤料深度反硝化性能研究[D]. 北京: 北京工业大学, 2013. [57] PAN X, GU Z, CHEN W, et al. Preparation of biochar and biochar composites and their application in a Fenton-like process for wastewater decontamination: A review[J]. Science of the Total Environment, 2021, 754: 142104. doi: 10.1016/j.scitotenv.2020.142104 [58] 王宁, 黄磊, 罗星, 等. 生物炭添加对曝气人工湿地脱氮及氧化亚氮释放的影响[J]. 环境科学, 2018, 39(10): 4505-4511. [59] 刘杰, 韩士群, 齐建华, 等. 生物碳含量对底泥活化原位脱氮及微生物活性的影响[J]. 江苏农业学报, 2016, 32(1): 106-110. doi: 10.3969/j.issn.1000-4440.2016.01.016 [60] 杨磊, 郭军, 王岩, 等. 农林废弃物制备环境友好型缓释碳源生物膜载体材料[J]. 湖北农业科学, 2021, 60(18): 68-71. [61] 陈佳伟, 许晓毅, 时和敏, 等. 基于D-最优混料设计的复合缓释碳源填料制备及其性能分析[J]. 环境污染与防治, 2021, 43(6): 718-724. [62] FALLAHI A, REZVANI F, ASGHARNEJAD H, et al. Interactions of microalgae-bacteria consortia for nutrient removal from wastewater: A review[J]. Chemosphere, 2021, 272: 129878. doi: 10.1016/j.chemosphere.2021.129878 [63] 丁绍兰, 谢林花, 马蕊婷. 壳类生物质释碳性能研究[J]. 环境污染与防治, 2016, 38(10): 1-5,11. [64] 朱辉翔, 张树楠, 彭英湘, 等. 不同固体碳源释碳特征及其对反硝化脱氮效果研究[J]. 农业现代化研究, 2021, 42(2): 206-214. [65] MULLER W R, HEINEMANN A, SCHAFER C, et al. Aspects of PHA(poly-B-hydroxy-butyric-acid) as an H-donator for denitrification in water-treatment processes// Int water supply assoc[J]. International workshop on inorganic nitrogen compounds and water supply in 1991. Hamburg, 1992: 27-29. [66] BOERSMA M, ABERLE N, HANTZSCHE F M, et al. Nutritional limitation travels up the food chain[J]. International Review of Hydrobiology, 2008, 93(4/5): 479-488. [67] STUTTER M I, GRAEBER D, EVANS C D, et al. Balancing macronutrient stoichiometry to alleviate eutrophication[J]. Science of the Total Environment, 2018, 634: 439-447. doi: 10.1016/j.scitotenv.2018.03.298 [68] VASSILEV S V, BAXTER D, ANDERSEN L K, et al. An overview of the organic and inorganic phase composition of biomass[J]. Fuel, 2012, 94(1): 1-33. [69] GILBERT P M. Eutrophication, harmful algae and biodiversity - Challenging paradigms in a world of complex nutrient changes[J]. Marine Pollution Bulletin, 2017, 124(2): 591-606. doi: 10.1016/j.marpolbul.2017.04.027 [70] REDFIELD A C. The biological control of chemical factors in the environment[J]. Science progress, 1960, 11: 150-170. [71] HILLEBRAND H, STEINERT G, BOERSMA M, et al. Goldman revisited: Faster-growing phytoplankton has lower N: P and lower stoichiometric flexibility[J]. Limnology and Oceanography, 2013, 58(6): 2076-2088. doi: 10.4319/lo.2013.58.6.2076 [72] 于潘, 张黎烜, 尤庆敏, 等. 综合硅藻指数的建立及其在淡水生态评价中的应用[J]. 环境科学研究, 2022, 35(9): 2165-2174. doi: 10.13198/j.issn.1001-6929.2022.06.03 [73] ISHIDA C K, ARNON S, PETERSON C G, et al. Influence of algal community structure on denitrification rates in periphyton cultivated on artificial substrata[J]. Microbial Ecology, 2008, 56(1): 140-152. doi: 10.1007/s00248-007-9332-0 -
 点击查看大图
点击查看大图
计量
- 文章访问数: 6247
- HTML全文浏览数: 6247
- PDF下载数: 165
- 施引文献: 0





















































