

-
研发可促进反硝化效果的生物强化技术是提升自然水体反硝化脱氮效能的关键,其中提供额外的无机电子供体是为反硝化细菌提供外部电子供体的最佳选择[1]。纳米零价铁 (nZVI) 作为一种环境友好的电子供体,已在废水处理、土壤修复等领域被用于促进反硝化脱氮[2]。但在铁颗粒微细化过程中,材料表面活性和自由能增强,颗粒表面原子极易与其他原子发生反应,导致表面钝化。此外,颗粒间团聚也会降低材料的反应性,限制了nZVI在实际修复领域中的发展。减少nZVI聚集的一种重要方法是将其负载到载体材料上,可用载体包括活性炭 (碳铁) [3]、碳纳米管[4]、石墨烯[5]、生物炭[6]、层状双氢氧化物[7]等。
此外,将nZVI与反硝化细菌结合可能是提高电子利用效率、提升反硝化速率、增强环境修复效果的有效途径。已有研究将反硝化菌与nZVI进行耦合,并开展了系列间歇实验考察系统反硝化脱氮效率。结果证实Fe0促进了硝酸盐的电子转移和厌氧生物转化[8]。因此,推测nZVI与反硝化菌等微生物的耦合不仅可以为微生物提供能量以满足其生长和代谢需求,还可以提高环境中的生物利用度[9]。
在众多nZVI负载材料中,具有高比表面积的生物炭兼具优越的材料性能和生物相容性, ZVI-BC-微生物体系不仅可以提高微生物反硝化效率,还可以提高电子传输速率,因此复合材料生物炭负载纳米零价铁 (BC/ZVI) 已在各类污染控制和环境修复过程中得以应用[10]。生物炭可以作为Pseudomonas citronellolis等微生物的庇护所,增强其对高浓度苯酚的生物降解效率,同时提供一定的可生物降解碳源[11]。ZHANG等[12]研究发现生物炭作为电子穿梭和导电材料,通过介导细胞外电子转移直接促进微生物代谢,从而促进硝酸盐的微生物还原。代谢组学能够定量监测反硝化菌代谢产物中的低分子量化合物[13],代谢物质的变化可用来评估微生物对不同材料添加的响应[14]。对代谢物质的相关分析可以深入了解微生物的代谢通路,揭示生物炭负载零价铁 (BC/ZVI) 强化脱氮副球菌去除硝酸盐的作用机制。
为探究单一反硝化功能菌在BC/ZVI-微生物体系中对硝酸盐去除效率和去除机制的影响,本研究选取反硝化模式微生物——脱氮副球菌 (Paracoccus denitrifican BNCC336866) 开展批次实验,通过分析BC、ZVI和BC/ZVI 3种材料对脱氮副球菌反硝化脱氮效率的影响,考察BC/ZVI强化脱氮副球菌硝酸盐去除过程,并借助非靶向代谢组学分析不同实验条件下反硝化过程中脱氮副球菌代谢通路发生的变化。
-
以干燥杨木原料过40目筛,在氮气氛围下 (120 mL·min−1) 升温至500 ℃,热解2 h获得原始生物炭。将1.0 g 原始生物炭与100 mL 1 M HCl溶液混合,搅拌24 h后,用去离子水洗2~3遍,真空泵进行抽滤,使出水pH 值达到6.0左右。剩余生物炭颗粒在真空干燥箱中干燥24 h,获得样品定义为BC。配置100 mL甲醇-水混合液 (3∶7,v/v) ,通氮气15 min,将 8.0 g的FeSO4·7H2O溶于甲醇-水混合液中,继续通氮气5 min,密封,而后在N2气氛下滴加0.5 M的NaOH,使溶液pH达到6.1,接着将100 mL含7.6 g NaBH4的溶液滴加到混合物中,在氮气氛围下搅拌1 h,后用甲醇洗2~3次,于真空干燥箱中干燥,获得的样品定义为ZVI。将8.0 g FeSO4·7H2O溶于甲醇-水混合液,通氮气5 min,密封,使其完全溶解。之后加入3.2 g BC,通氮气10 min,密封反应6 h,后续操作同ZVI制备过程,洗涤干燥后所得样品定义为BC/ZVI。
-
采用X射线光电子能谱 (XPS) 分析样品表面组成。采用BET方法测定样品的比表面积、孔体积和孔径分布,采用FTIR测定表面官能团组成,扫描范围为4 000~400 cm−1。采用循环伏安法 (CV) 和电化学阻抗谱法 (EIS) 对材料的电化学行为进行分析,以在ITO板上制备的材料电极为工作电极,氯化银电极为参比电极,铂网电极为对电极构成的三电级系统对每组材料进行检测。
-
脱氮副球菌Paracoccus denitrifican购买自北纳菌种保藏中心,菌种编号为BNCC336866。将实验器具和LB培养基于121 ℃条件下灭菌15 min。将菌株接种到新鲜的LB培养基中,在30 ℃、150 r·min−1摇床培养24 h后得到菌液。接着将菌液转移至50 mL离心管中,7 000 r·min−1条件下离心15 min,去除上清液,沉淀的菌体用无菌水洗涤离心,重复3次。加入适量无菌水将沉淀的菌体稀释成OD600为0.7的菌悬液备用。
-
使用含有50 mL反硝化培养基 (pH=7.0) 的系列100 mL血清瓶进行批次实验。将反硝化培养基在121 ℃高压灭菌15 min,冷却至室温后使用。实验过程中保证各类营养素的外源供应超过微生物生长需求,以降低从生物炭中浸出的部分营养物质对细菌生长的影响。培养溶液中加入50 mmol·L−1 HEPES缓冲溶液,避免由于材料添加引起的pH变化。
在反硝化介质中分别加入不同含量的BC、ZVI及BC/ZVI,用0.1 mol·L−1 NaOH或HCl 将每个血清瓶的pH调节到7.0,用纯Ar除氧后,将DO浓度维持在0.5 mg·L−1以下,保持厌氧状态。向灭菌后的血清瓶中接种P.denitrificans,将接种后细胞光密度 (OD600) 控制在0.02,以确保相近的初始细胞密度 (2×107细胞·mL−1) ,随后置于摇床 (150 r·min−1,30 ℃) 中于暗处培养。同时设置非生物处理组,如表1所示。在培养开始后0、6、12、18和24 h使用无菌注射器分别从各样品组中提取2 mL溶液,用孔径为0.22 μm滤膜对溶液进行过滤,滤液储存在4 ℃条件下,采用标准方法检测滤液中硝酸盐含量。
-
基于液质联用 (LC-MS) 技术分别对仅添加微生物组和BC/ZVI高添加量组、ZVI-高添加组和BC/ZVI-高添加量组、BC-高添加量组和BC/ZVI-高添加量组进行非靶向代谢组学研究。配置甲醇-水混合液 (4∶1,v/v) 用于提取细胞中的代谢物。取1 mL样本于冻干机中冻干,加入浓度为100 μL的甲醇水溶液。而后,涡旋震荡,冰浴中静置5 min,在4 ℃下以15 000 g离心15 min后,取一定量的上清液加质谱级纯水稀释至甲醇含量为53%。再次离心,收集上清液,进行LC-MS分析。采用UPLCHSST3柱 (2.1 mm×100 mm、1.8 μm) 、Q Exactive质谱仪 (安捷伦技术有限公司
1290 ,美国) 对样品进行LC-MS/MS测定。洗脱梯度设定为 0 min,1% B;1 min,1% B;8 min,99% B;10 min,99% B;10.1 min,1% B;12 min,1% B。每次进样体积2 μL。Q Exactive质谱仪在Xcalibur软件控制下进行一级、二级质谱数据采集。 -
材料的红外光谱数据表明BC和BC/ZVI的表面由丰富的含氧官能团组成。2 825 cm−1附近的弱峰分别来自脂肪族官能团 (-CH3、-CH2-) 的不对称和对称拉伸,在1 581 cm−1附近的峰表明共轭酮和醌上羰基基团的存在,盐酸改性后,BC在1 581 cm−1处的吸收增强[15] 。研究表明,酚羟基和醌羰基基团同时参与给电子和受电子的过程,在硝酸盐反硝化过程中起关键作用。1 155 cm−1左右的吸收峰代表脂肪性C-O-C的振动吸收,BC/ZVI 的C-O-C峰强度远高于BC,很可能是铁可以与生物质中的氧原子发生反应,从而增加生物炭上表面官能团的含量[16]。
图1(a)表明ZVI成功负载在生物炭上,C 1s分谱图(图1(b))中284.8 eV的主峰对应于sp2 杂化碳(C-C/C=C),而sp3杂化碳 285.6~286.2 eV和289.0~289.4 eV附近的峰值分别属于C-O和O-C=O [17]。O 1s谱图 (图1(c)) 中观察到的532.6~532.7 eV附近的峰可分别归因于O-C=O,533.7 eV附近的峰属于C=O。BC谱图中,在531.96 eV附近观察到C-O的存在,而在BC/ZVI中C-O消失,却在531.08 eV处观察到了Fe-OH的存在。对BC/ZVI (图1(d)) 的Fe 2p谱进行拟合,707 eV处的强峰证实了Fe0的成功合成。同时,在709.3 eV (2p1/2)和722.9 eV (2p1/2)处的双峰表明表面存在Fe (II) ,Fe 2p3/2 (726.4 eV) 和Fe 2p3/2 (711.8 eV) 的宽峰代表了Fe (Ⅲ) 的形成[18]。这些发现表明,BC/ZVI表面的钝化层可分为2类,即Fe3O4/FeO、Fe2O3/FeOOH。
-
图2(a)显示3种材料的CV曲线,其中BC/ZVI的面积最大,其次是BC、ZVI,这证实零价铁的负载显著增加了材料的电容。在-0.5V附近具有更正的氧化峰,表明BC/ZVI比ZVI和BC产生了更多的氧化还原物种。当单独使用BC或ZVI作为工作电极、KCl (0.1mol·L−1) 作为电解质时,得到的CV曲线中未观察到明显的氧化还原峰。相反,在BC/ZVI上观察到明显的氧化还原电流,这表明ZVI可能成为系统的电子中心。采用EIS曲线的半圆直径来评估材料和溶液之间直接电子转移的难易程度[19]。与BC和ZVI相比,BC/ZVI材料具有更小的半圆直径,表明BC/ZVI具有最低的电荷转移电阻 (Rct) 和最强的电子转移能力。经Zview软件拟合,BC、ZVI和BC/ZVI的Rct分别为19.4、15.8和14.2 Ω,说明BC/ZVI具有最低的阻抗。在低频区,BC/ZVI的Nyquist曲线斜率明显大于BC和ZVI,更接近于垂直,而在高频处BC/ZVI的Z″值更低,表明其具有更好的电子传输能力[20-21]。
-
首先考察了不同处理体系中吸附和还原作用对硝酸盐去除的具体贡献比例。对于仅投加BC的体系,实验进行24 h后低添加量和高添加量组对硝酸盐的吸附分别为3.0%±0.2%和5.1%±0.3% (图3(a)) 。研究证实底物和细胞间的传质对于总生物降解效率至关重要,而生物炭可以有效地吸附硝酸盐和有机物,并为微生物释放营养物质。通过比较低添加量ZVI组和高添加量ZVI组的差异可以发现ZVI的还原作用可贡献总硝酸盐去除的5.3%±0.8%和15.6%±0.7% (图3(a)) ,这种较弱的硝酸盐还原能力可以归因于ZVI的腐蚀。
然后进行批次实验确定实验所需菌液投加量,分别在OD600=0.02、0.03、0.04、0.05和0.1的细菌投加量下测定了相应的硝酸盐去除率 (图3(b)) 。结果表明当OD600=0.02时,24 h内硝酸盐的去除率为70%±3.79%。而更高投加量下,20 h内硝酸盐去除率均在100%附近。为了后期实验的对比差异,选取OD600=0.02为后期实验所需菌液浓度。
如图3(c)所示,BC+反硝化菌体系、ZVI+反硝化菌体系及BC/ZVI+反硝化菌体系中硝酸盐去除效果均优于仅含有反硝化菌的体系。相同投加量条件下,硝酸盐去除效果为BC/ZVI+反硝化菌体系>ZVI+反硝化菌体系>BC+反硝化菌体系>反硝化菌体系。具体而言,在仅含有反硝化菌的体系中,反硝化作用在4 h的稳定期后开始,24 h后可除去体系中70.0%±3.8%的硝酸盐。与纯细菌处理体系相比,添加功能材料 (BC、ZVI和BC/ZVI) 的体系中硝酸盐均发生快速去除,几乎没有停滞期。实验进行24 h后,由于碳源的缺乏,仅添加反硝化菌体系中硝酸盐去除率不再上升。而对于添加生物炭的实验组,实验进行24 h时,低添加量和高添加量BC组中硝酸盐去除率可进一步提升至73.4%±1.7%和77.9%±1.4%。这可能是由于生物炭能够为脱氮副球菌提供一定量的碳源并且含有酚羟基等氧化还原官能团 (图1) 。
添加ZVI后,体系中存在的氧化还原反应Fe0→Fe2+→Fe3+会促进系统间电子转移过程,加速体系硝酸盐还原进程,反应16 h后低添加量和高添加量ZVI组中NO3−去除率从43.3%提高到77.7%±2.2%和65.4%±2.2% (16 h) ,这可能是由于ZVI材料较BC具有更低的阻抗和更优越电子传输能力 (图2) 。较高的铁浓度已被证明可以促进细菌代谢,提高微生物活性,上调反硝化基因转录以促进硝酸盐还原[22]。在BC/ZVI+反硝化菌体系中,低添加量和高添加量组中24 h内实现100%和92.7%±3.3%的硝酸盐去除,而在只添加BC/ZVI的非生物体系中,低添加量和高添加量组中只有21.5%±1.5%和42.2%±1.4%的硝酸盐被去除。研究发现,在添加Fe (0) 的体系中,厌氧铁腐蚀过程产生的Fe (II) 可能作为硝酸盐的电子供体,证明了使用nZVI铁作为生物硝酸盐还原电子源的潜在适用性[2]。
-
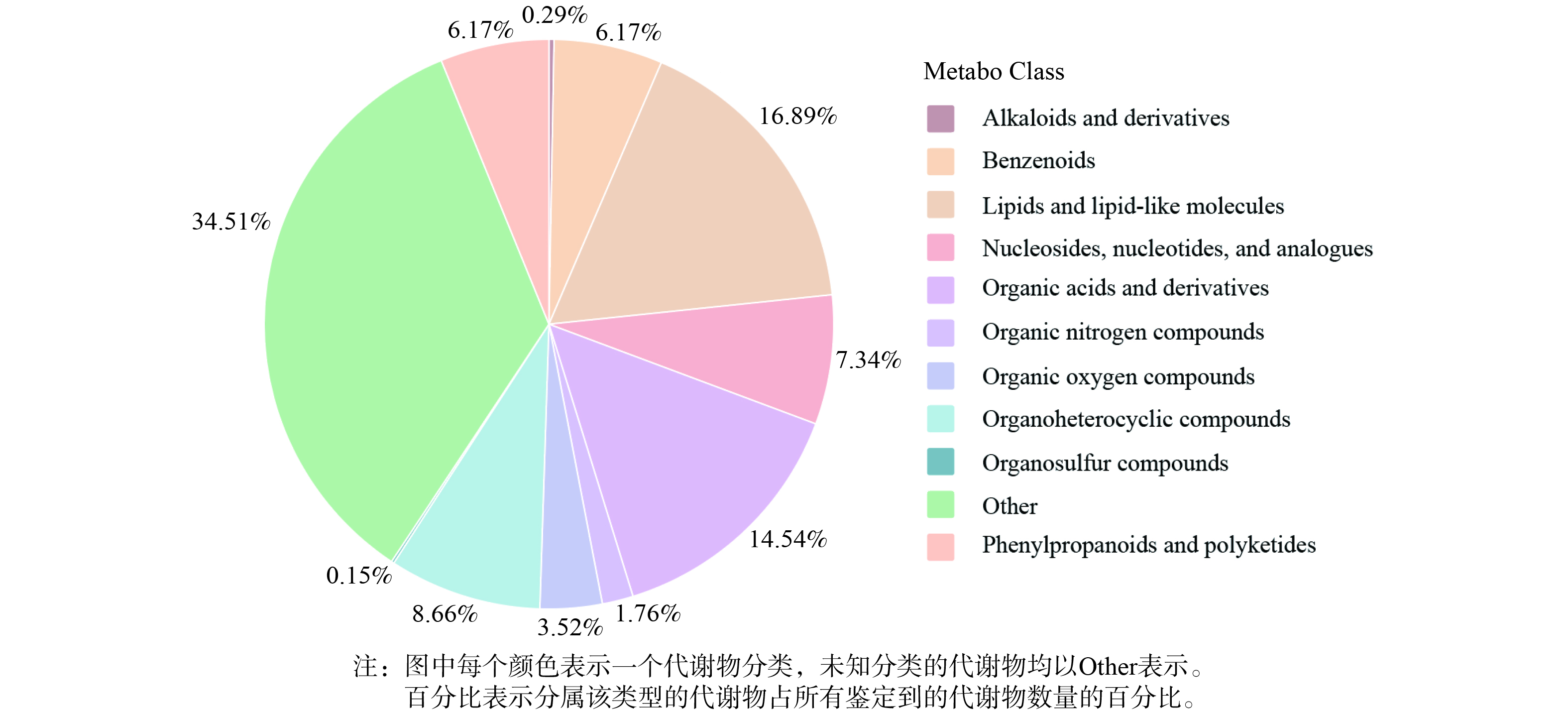
1) 代谢物分类。将所有实验组鉴定到的代谢物根据化学分类归属信息进行分类统计,各类代谢物占比如图4所示。代表性代谢物及占比如下:生物碱及其衍生物 (alkaloids and derivatives,0.29%) 、苯环型 (benzenoids,6.17%) 、脂类和类脂类分子 (lipids and lopid-like molecules,16.89%) 、核苷、核苷酸及其类似物 (nucleosides,nucleotides,and analogues,7.34%) 、有机酸及其衍生物 (organic acids and derivatives,14.54%) 、有机氮化合物 (organic nitrogen compounds,1.76%) 、有机氧化合物 (organic oxygen compounds,3.52%) 、有机杂环化合物 (organoheterocyclic compounds,8.66%) 、有机硫化合物 (organosulfur compounds,0.15%) 、苯丙类和聚酮类 (phenylpropanoids and polyketides,6.17%) 等。
2) 代谢物和差异代谢物的筛选及分析。经过对两组数据的整理,使用CD3.1数据处理软件,对样本中检测到的色谱峰进行积分。其中每个特征峰的峰面积表示一个代谢物的相对定量值,同时用blank样本去除背景离子,并使用总峰面积对原始定量结果进行标准化处理。然后保留QC样本中变异系数 (coefficient of variance,CV) 小于30%的代谢物,最终得到代谢物的鉴定和相对定量结果。根据OPLS-DA模型,得到代谢物的变量投影重要度VIP,排序筛选出VIP>1的差异代谢物并结合单变量分析的差异倍数值 (fold change) 筛选差异表达代谢物。本文分别将仅添加微生物组和BC/ZVI-高添加量组、ZVI-高添加量组和BC/ZVI-高添加量组、BC-高添加量组和BC/ZVI-高添加量组进行处理,通过火山图 (volcano plot) 对差异表达代谢物进行可视化。
由图5(a)可知共筛选到93种具有统计学显著差异的代谢物。与仅添加微生物组相比,BC/ZVI-高添加量组中微生物的代谢产物有19种代谢物表达下调,74种代谢物表达上调。添加BC/ZVI后,丰度显著上调的代谢物数量远高于丰度显著下调的代谢物数量,由此初步表明BC/ZVI的添加对微生物代谢具有促进作用。由图5(b)可知共筛选到60种具有统计学显著差异的代谢物。与ZVI-高添加量组对比,BC/ZVI-高添加量组中微生物的代谢产物有1种代谢物表达下调,59种代谢物表达上调,这可能是因为BC/ZVI材料表面形成钝化层有效减少了nZVI颗粒团聚,提升了材料与微生物的有效接触面积,强化了材料与微生物间的相互作用。BC/ZVI-高添加量组丰度显著上调的代谢物数量远高于丰度显著下调的代谢物数量,由此初步表明对于添加BC后的ZVI组对微生物代谢更具有促进作用。由图5(c)可知此时共筛选136种具有统计学显著差异的代谢物。与BC-高添加量组相比,BC/ZVI-高添加量组中微生物的代谢产物有102中代谢物下调,34种代谢物上调。由此可初步推断nZVI生物毒性在一定程度上会抑制微生物代谢,但是后续还要对上调的差异代谢物进行代谢富集通路分析。
3) 上调差异代谢物的KEGG富集通路分析。图6(a)表明,BC/ZVI添加后脱氮副球菌代谢产物中上调的代谢物与多类功能有关,包括ABC转运蛋白、柠檬酸循环/三羧酸循环、鸟氨酸,赖氨酸和烟酸衍生的生物碱的生物合成、酪氨酸代谢、丙氨酸,天冬氨酸和谷氨酸代谢、双组分系统、萜类和类固醇的生物合成、嘧啶代谢、核苷酸代谢、原核生物的固碳途径、苯丙氨酸代谢、辅助因子的生物合成、组氨酸和嘌呤衍生的生物碱的生物合成、嘌呤代谢和碳代谢等。涉及到碳水化合物代谢 (与碳氧化有关,如三羧酸循环) 、能量代谢 (与氮还原有关,如原核生物的固碳途径) 以及与细胞生长相关的途径 (如各种氨基酸代谢) 。图6(b)表明,上调差异代谢物通路主要与代谢途径、柠檬酸循环/三羧酸循环、微生物在不同环境中的代谢、嘌呤代谢、丙氨酸,天冬氨酸和谷氨酸代谢、次生代谢产物的生物合成、双组分系统、嘧啶代谢、酪氨酸代谢、鸟氨酸,赖氨酸和烟酸衍生的生物碱的生物合成等相关。图6(c)表明,上调差异代谢物代谢通路主要与代谢途径、微生物在不同环境中的代谢、次生代谢产物的生物合成、鸟氨酸,赖氨酸和烟酸衍生的生物碱的生物合成、苯丙烷类化合物的生物合成、氨基苯甲酸脂降解、谷胱甘肽代谢有关。
氨基酸是生物体的组成部分,在物质的合成中起着至关重要的作用。酪氨酸是所有生物体蛋白质合成所需的芳香族氨基酸,但仅在植物和微生物体[23]内合成。赖氨酸在蛋白质生成中起重要作用,它能够形成酰胺键成为翻译后修饰的位点,包括乙酰化、甲基化和泛素化。细胞对应激的反应需要由这些修饰引起的蛋白质功能和细胞形态的变化[24]。因此,在氧化应激的反应中,含有赖氨酸的蛋白质和代谢物可能起关键作用。此外,线粒体电子传递链与氨基酸分解代谢和赖氨酸的三羧酸循环有关。丙氨酸是蛋白质和细胞壁肽聚糖的重要组成部分,天冬氨酸和谷氨酸是合成各种氨基酸和碱的重要氮供体[25]。以上代谢产物的丰度上调表明,BC/ZVI添加增强了脱氮副球菌对多种物质的转化过程。
ABC转运蛋白在生物体内承担着多种物质跨膜运输的作用。一方面,可以实现多种营养物质从胞外向细胞质基质的转运,促进细胞的生长;另一方面,能够将多种不利物质通过外转运排出胞外,有利于将胞内非必须的次级代谢物或外源物质维持在较低浓度水平,减轻细胞压力,有助于细胞的生存与生长[26]。通过对比发现添加BC/ZVI使参与ABC转运的去氧胞苷作用上调,这说明BC/ZVI的添加促进了脱氮副球菌的跨膜转运,促进细胞的生长。TCA循环是碳水化合物、脂肪和氨基酸的最终常见氧化途径,它是向身体提供能量的最重要的代谢途径,是连接个体代谢途径的最重要的中枢途径。相较于仅添加微生物组,在BC/ZVI添加的体系中,柠檬酸和延胡索酸这两种参与TCA循环的代谢物出现上调,这说明BC/ZVI添加促进了脱氮副球菌的能量供应和物质代谢。
-
1) 材料表征分析证实BC/ZVI上的C-O-C峰强度远高于BC,很可能是铁可以与生物质中的氧原子发生反应,从而增加生物炭上表面官能团的含量。BC/ZVI表面的钝化层可分为两类,即Fe3O4/FeO、Fe2O3/FeOOH,可能是BC/ZVI材料强化脱氮副球菌去除硝酸盐的原因之一。EIS结果表明,与BC和ZVI相比,BC/ZVI复合材料具有最低的阻抗和最强的电子转移能力。不同材料的阻抗顺序为BC/ZVI<ZVI<BC。
2) 通过批次实验考察了BC、ZVI、BC/ZVI存在下硝酸盐的去除率。反应24 h后,生物炭对硝酸盐的吸附去除量为3%~5%,ZVI的还原作用可贡献硝酸盐总去除量的5.3%~15.6%。相较于仅添加脱氮副球菌的对照组,BC、ZVI和BC/ZVI的添加不同程度上强化了体系的脱氮效能。相同投加量条件下,硝酸盐去除效果为BC/ZVI+脱氮副球菌体系>ZVI+脱氮副球菌体系>BC+脱氮副球菌体系>仅添加脱氮副球菌体系。
3) 通过非靶向代谢组学分析证实相较于仅添加微生物和ZVI高添加量的实验组,BC/ZVI高添加量组丰度显著上调的代谢物数量远高于丰度显著下调的代谢物数量。这些上调的代谢物主要涉及ABC转运蛋白、三羧酸循环、丙氨酸,天冬氨酸和谷氨酸代谢等代谢通路。这说明BC/ZVI主要从促进脱氮副球菌的跨膜转运过程、物质代谢和能量供应、蛋白质和细胞壁的合成等方面增强脱氮副球菌细胞生长与增殖,从而强化其脱氮过程。
生物炭负载纳米零价铁强化脱氮副球菌去除硝酸盐及微生物代谢特征解析
nZVI materials loaded on biochar enhance the nitrate removal by denitrifying bacteria and the microbial metabolic characteristics
-
摘要: 纳米零价铁 (nZVI) 与反硝化菌等微生物的耦合在河湖硝酸盐修复领域具有应用潜力,但存在nZVI生物毒性较强、颗粒易团聚等问题,将其负载到载体材料上可以有效缓解以上问题。其中生物炭负载nZVI材料在促进硝酸盐去除方面表现出优越性能,但现阶段该材料强化反硝化菌生物脱氮的机制尚不明确。本研究制备了代表性生物炭负载纳米零价铁材料 (BC/ZVI) ,接着采用反硝化模式微生物脱氮副球菌 (Paracoccus denitrifican BNCC336866) 为实验菌种,构建系列批次实验考察BC/ZVI强化脱氮副球菌硝酸盐去除过程,结果表明ZVI的还原作用可贡献硝酸盐总去除量的5.3%~15.6%。相较于仅添加脱氮副球菌的对照组,BC、ZVI和BC/ZVI的添加不同程度上强化了体系的脱氮效能。最后通过非靶向代谢组学分析证实BC/ZVI主要从促进脱氮副球菌的跨膜转运过程、物质代谢和能量供应、蛋白质和细胞壁的合成等方面增强脱氮副球菌细胞生长与增殖,从而强化其脱氮过程。
-
关键词:
- 生物炭负载纳米零价铁 /
- 反硝化 /
- 脱氮副球菌 /
- 代谢组学
Abstract: The coupling of nano-sized zero valent iron (nZVI) with microbes such as denitrifying bacteria has application potential in the field of nitrate remediation. However, several problems still remained during the applications, including the biological toxicity to microbes and easy aggregation of nZVI particles. Loading nZVI onto carrier materials can effectively alleviate these problems. Among various loading materials, nZVI materials loaded on biochar exhibit superior performance in promoting denitrification efficiency. However, the mechanism by which biochar loaded nZVI enhances denitrification by denitrifying bacteria is still unclear. This study synthesized representative nZVI materials loaded on biochar (BC/ZVI). Paracoccus denitrifican BNCC336866 was used as a model denitrification mode strain and a series of batch experiments was conducted to investigate the nitrate removal process in the presence of BC/ZVI. The results indicated that the reduction effect of ZVI can contribute 5.3% to 15.6% of the total nitrate removal. Compared to the control group that only dosed with denitrifying bacteria, the addition of BC, ZVI and BC/ZVI could enhance the denitrification efficiency to varying degrees. The untargeted metabolomics analysis confirmed that BC/ZVI predominantly enhanced the growth and proliferation of denitrifying bacteria cells by promoting transmembrane transport, substance metabolism and energy supply, protein and cell wall synthesis, etc., thereby strengthening the overall denitrification capacity. -

-

图 2 BC、ZVI和BC/ZVI的电化学性能解析
Figure 2. Electrochemical performance analysis of BC, ZVI and BC/ZVI
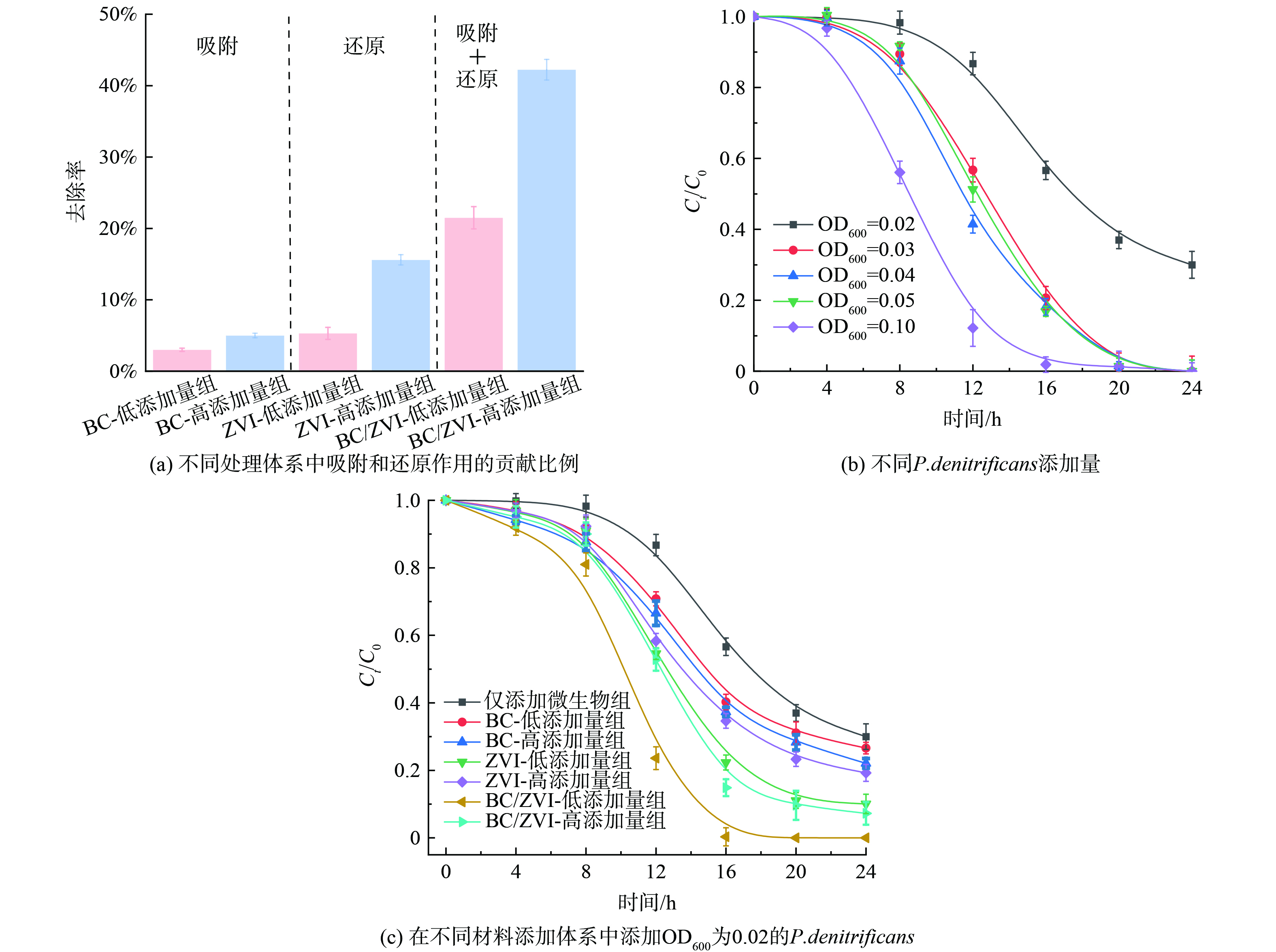
图 3 不同体系中硝酸盐去除效果
Figure 3. Removal efficiency of nitrate in variable treatment systems
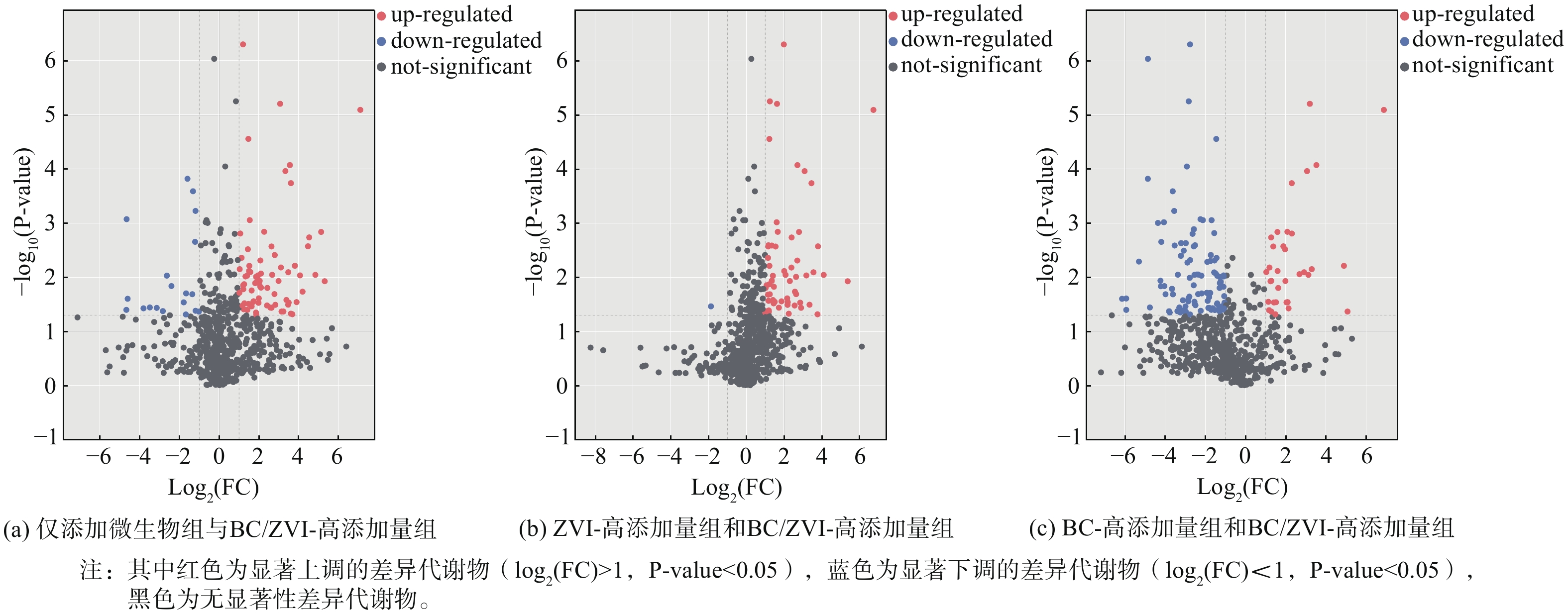
图 5 不同实验条件下差异表达代谢物火山图
Figure 5. Volcanic plots of differentially expressed metabolites in variable treatments

图 6 不同实验条件下上调差异表达代谢物KEGG通路分析
Figure 6. KEGG pathway analysis of up-regulated differentially expressed metabolites in variable treatments
表 1 批次实验设置
Table 1. Setting up of batch experiments
物质添加情况 无菌无材料添加组 BC-低添加量 BC-高添加量 ZVI-低添加量 ZVI-高添加量 BC/ZVI-低添加量 BC/ZVI-高添加量 微生物/OD值 0 0 0 0 0 0 0 材料/ (g·L−1) 0 0.2 0.4 0.1 0.2 0.3 0.6 硝酸盐/ (mg·L−1) 30 物质添加情况 仅添加微生物组 BC-低添加量 BC-高添加量 ZVI-低添加量 ZVI-高添加量 BC/ZVI-低添加量 BC/ZVI-高添加量 微生物/OD值 0.02 0.02 0.02 0.02 0.02 0.02 0.02 材料/ (g·L−1) 0 0.2 0.4 0.1 0.2 0.3 0.6 硝酸盐/ (mg·L−1) 30  下载: 导出CSV
下载: 导出CSV
-
[1] JIANG M, ZHENG X, CHEN Y. Enhancement of denitrification performance with reduction of nitrite accumulation and N2O emission by Shewanella oneidensis MR-1 in microbial denitrifying process[J]. Water Research, 2020, 169: 115242. doi: 10.1016/j.watres.2019.115242 [2] SHIN K H, CHA D K. Microbial reduction of nitrate in the presence of nanoscale zero-valent iron[J]. Chemosphere, 2008, 72(2): 257-262. doi: 10.1016/j.chemosphere.2008.01.043 [3] MACKENZIE K, BLEYL S, GEORGI A, et al. Carbo-Iron-An Fe/AC composite-as alternative to nano-iron for groundwater treatment[J]. Water Research, 2012, 46(12): 3817-3826. doi: 10.1016/j.watres.2012.04.013 [4] XU J, LV X, LI J, et al. Simultaneous adsorption and dechlorination of 2, 4-dichlorophenol by Pd/Fe nanoparticles with multi-walled carbon nanotube support[J]. Journal of Hazardous Materials, 2012, 225: 36-45. [5] SUN Y, DING C, CHENG W, et al. Simultaneous adsorption and reduction of U(VI) on reduced graphene oxide-supported nanoscale zerovalent iron[J]. Journal of Hazardous Materials, 2014, 280: 399-408. doi: 10.1016/j.jhazmat.2014.08.023 [6] SU H, FANG Z, TSANG P E, et al. Stabilisation of nanoscale zero-valent iron with biochar for enhanced transport and in-situ remediation of hexavalent chromium in soil[J]. Environmental Pollution, 2016, 214: 94-100. doi: 10.1016/j.envpol.2016.03.072 [7] SHENG G, TANG Y, LINGHU W, et al. Enhanced immobilization of ReO4− by nanoscale zerovalent iron supported on layered double hydroxide via an advanced XAFS approach: Implications for TcO4− sequestration[J]. Applied Catalysis B-Environmental, 2016, 192: 268-276. doi: 10.1016/j.apcatb.2016.04.001 [8] OH S Y, SEO Y D, KIM B, et al. Microbial reduction of nitrate in the presence of zero-valent iron and biochar[J]. Bioresource Technology, 2016, 200: 891-896. doi: 10.1016/j.biortech.2015.11.021 [9] 代快, 李江舟, 蒲天燕, 等. 施用生物炭对3种烟用农药残留的影响[J]. 中国农业科技导报, 2019, 21(8): 99-106. [10] YU T, WANG L, MA F, et al. A bio-functions integration microcosm: Self-immobilized biochar-pellets combined with two strains of bacteria to remove atrazine in water and mechanisms[J]. Journal of Hazardous Materials, 2020, 384: 121326. doi: 10.1016/j.jhazmat.2019.121326 [11] ZHAO L, XIAO D, LIU Y, et al. Biochar as simultaneous shelter, adsorbent, pH buffer, and substrate of Pseudomonas citronellolis to promote biodegradation of high concentrations of phenol in wastewater[J]. Water Research, 2020, 172: 115494. doi: 10.1016/j.watres.2020.115494 [12] ZHANG Y, ZHANG Z, CHEN Y. Biochar mitigates N2O emission of microbial denitrification through modulating carbon metabolism and allocation of reducing power[J]. Environmental Science & Technology, 2021, 55(12): 8068-8078. [13] QI X, YIN H, ZHU M, et al. Understanding the role of biochar in affecting BDE-47 biodegradation by Pseudomonas plecoglossicida: An integrated analysis using chemical, biological, and metabolomic approaches[J]. Water Research, 2022, 220: 118679. doi: 10.1016/j.watres.2022.118679 [14] 陈慧, 马徐, 王海波, 等. 供水PE和PPR塑料管内表面生物膜群落组成及其代谢特征[J]. 环境科学学报, 2023, 43(12): 287-295. [15] LI B, YANG L, WANG C Q, et al. Adsorption of Cd(II) from aqueous solutions by rape straw biochar derived from different modification processes[J]. Chemosphere, 2017, 175: 332-340. doi: 10.1016/j.chemosphere.2017.02.061 [16] THI HANH N, THI HUONG P, HONG THAM NGUYEN T, et al. Synthesis of iron-modified biochar derived from rice straw and its application to Arsenic Removal[J]. Journal of Chemistry, 2019, 2019: 1-8. [17] WAN Z, LI K. Effect of pre-pyrolysis mode on simultaneous introduction of nitrogen/oxygen-containing functional groups into the structure of bagasse-based mesoporous carbon and its influence on Cu(II) adsorption[J]. Chemosphere, 2018, 194: 370-380. doi: 10.1016/j.chemosphere.2017.11.181 [18] DAI X H, FAN H X, YI C Y, et al. Solvent-free synthesis of a 2D biochar stabilized nanoscale zerovalent iron composite for the oxidative degradation of organic pollutants[J]. Journal of Materials Chemistry A, 2019, 7(12): 6849-6858. doi: 10.1039/C8TA11661J [19] WU L, LIN Q, FU H, et al. Role of sulfide-modified nanoscale zero-valent iron on carbon nanotubes in nonradical activation of peroxydisulfate[J]. Journal of Hazardous Materials, 2022, 422: 124969. [20] YANCHUS D A, KIRK D W, JIA C Q. Investigating the effects of biochar electrode macrostructure and dimension on electrical double-layer capacitor performance[J]. Journal of the Electrochemical Society, 2018, 165(2): A305-A313. doi: 10.1149/2.1151802jes [21] 邓宇. 生物炭基材料的制备及其电化学性能研究[D]. 长沙: 湘潭大学, 2020. [22] FENG L, YANG J, MA F, et al. Biological stimulation with Fe(III) promotes the growth and aerobic denitrification of Pseudomonas stutzeri T13[J]. Science of the Total Environment, 2021, 776: 145939. doi: 10.1016/j.scitotenv.2021.145939 [23] SCHENCK C A, MAEDA H A. Tyrosine biosynthesis, metabolism, and catabolism in plants[J]. Phytochemistry, 2018, 149: 82-102. doi: 10.1016/j.phytochem.2018.02.003 [24] WANG J, YIN J, PENG D, et al. 4-Nitrophenol at environmentally relevant concentrations mediates reproductive toxicity in Caenorhabditis elegans via metabolic disorders-induced estrogen signaling pathway[J]. Journal of Environmental Sciences, 2025, 147: 244-258. doi: 10.1016/j.jes.2023.09.032 [25] REITZER L. Biosynthesis of glutamate, aspartate, asparagine, l-alanine, and d-alanine[J]. EcoSal Plus, 2004, 1(1). [26] 曲俊泽, 陈天华, 姚明东, 等. ABC转运蛋白及其在合成生物学中的应用[J]. 生物工程学报, 2020, 36(9): 1754-1766. -
 点击查看大图
点击查看大图

计量
- 文章访问数: 1876
- HTML全文浏览数: 1876
- PDF下载数: 23
- 施引文献: 0
