

-
全球变暖已成事实,《中国气候变化蓝皮书》(2021)显示[1],包括中国在内的全球气候变暖仍在持续,我国仍是全球气候变化的敏感区和影响显著区,升温速率明显高于全球同期平均水平。 气候变化将导致区域生态稳定性降低,改变温度、光照、水资源等农业关键要素的时空分布格局,极易引发干旱、洪涝等气象灾害,对农业生产构成严重威胁[2]。特别是干旱、半干旱区,生态系统脆弱,对气候变化的响应更为直接和敏感[3],而我国有超一半的国土面积属于干旱、半干旱区,这意味着我们将同时面临气候变暖与干旱的双重威胁,由此可能对农业生产、环境生态、人类健康等诸多领域产生深远影响[4]。《The Lancet-Planetary Health》发布的研究显示[5],未来30年,气候变化将导致小麦、水稻、玉米等粮食作物中核心营养元素如蛋白质、铁和锌的可供应量将分别降低19.5%、14.4%和14.6%。上述问题通过食物链的逐级放大作用,势必造成全球范围内的“隐形饥饿”(营养素摄入不足或营养失衡)问题的进一步凸显[6],并最终可能威胁到人类的正常生存与发展。目前,国内外有关气候变暖、干旱等环境胁迫对作物矿质元素影响的研究已取得一定进展,如Li等[7]、Sardans等[8]的相关研究,但已有研究大多仅涉及单一因素,有关气候变暖与干旱复合胁迫的研究尚不多见,涉及亚细胞层面的探索则更为缺乏。
为全面深入探索气候变化对农业生态、食品品质、人类健康等可能产生的深远影响,揭示其规律及其内在的机理机制,本研究采用人工智能气候室,在干旱半干旱区,进行了气候变暖和干旱复合胁迫对小麦幼苗在亚细胞水平矿质元素动态富集的影响研究,旨在探索未来复杂气候变化背景下,作物矿质养分吸收、分布、利用的微观响应动态及其机理,以期为预估气候变化对农业生态、人类健康可能产生的远期效应提供理论依据,为农业系统科学应对气候变化提供重要参考。
-
本研究在中国气象局兰州干旱气象研究所试验基地( 35°35'N,104°37'E) 开展。基地气候主要特征为干旱少雨、光照强烈、无霜期长,年均温度为5.5—7.7 ℃,年均降雨量350—600 mm左右,属于半干旱地区气候环境。模拟实验在基地人工智能气候室进行,气候室的室内总面积594 m2,分割为互相独立的3个小气候室。气候室通过智能控制系统启闭相关子系统,实现对温度、湿度、光照和 CO2浓度等环境因子的自动控制,自运行以来已有多项研究在该气候室完成[9-10]。实验基地土壤以黄绵土为主,总氮为0.77 g·kg−1,碱解氮为79.73 mg·kg−1, 速效磷29.26 mg·kg−1,速效钾296.64 mg·kg−1,有机质含量为10.34 g·kg−1,pH 值为7.93。
-
气候变暖模拟分别在3个独立的气候室中进行,共设3个处理条件,如(表1), 实验设1 个对照处理组(A)和3个实验处理组(TD1、TD2、TD3),对照处理设在人工气候室外带移动式遮雨棚的自然田野,实验组分别设在上述3个独立的人工气候。每个处理区的大田均分为3个区,代表3次重复。选取优质西汉1号小麦种粒,经清洗消毒处理后,在湿润的石英砂中发芽,5 d后选取生长一致的小麦幼苗移栽至各处理田间土壤中,保持等行距20 cm。各处理土壤完全同质,施肥以底肥的方式一次性施肥,不再追肥,施肥标准统一为:有机肥 25 t·hm−2,磷酸二铵 250 kg·hm−2,尿素 150 kg·hm−2。每3 d仔细检查一次病虫害,实验期间未发生任何病虫害,未使用任何农药。生长35 d后,自各处理组的3个重复中采用梅花样布点的形式取5株整株样本,用去离子水洗净,根叶分离后置于-70 ℃冰箱保存,用于亚细胞矿质元素测定。
-
各处理组土壤在播种前自每个处理的各试验区内采用梅花样布点取5 个样品(25 cm×25 cm)。用塑料铲取0—20 cm耕作层土样,混匀后,称取约1 kg 样品,室内自然风干,105 ℃烘至恒重,充分研细后用100 目尼龙网过筛,4 ℃低温储存待测[7]。土壤水解氮的测定采用碱解扩散法,有效磷的测定采用碳酸氢钠提取-钼锑抗比色法,速效钾的测定采用乙酸铵提取-火焰光度法,有机质分析采用重铬酸钾容量法。Fe、Zn元素含量由兰州大学测试中心采用火焰原子吸收法分析。土壤Fe、Zn元素有效态采用DTPA 0.005 mol·L−1+CaCl2 0.01 mol·L−1+TEA 0.1 mol·L−1 (pH 7.3)溶液提取。
-
准确称取各处理小麦幼苗根/叶样品5 g,匀浆处理后采用低温差速离心技术进行亚细胞分级,将小麦幼苗根、叶分别分为F1( 细胞壁及细胞碎片) 、F2( 细胞器) 和F3( 细胞可溶性组分) 3 个亚细胞分级 [9]。Fe、Zn元素含量由兰州大学测试中心完成分析。
-
数据分析采用SPSS 22.0 for Windows ( SPSS Inc.,USA) 统计软件包进行.绘图采用Originpro 2016( OriginLab USA) 软件包进行。小麦对Fe、Zn元素的生物富集系数(BF)和转移系数(TF)的计算方法参见本课题组的前期研究[9]。
-
如表2、表3所示,播种前各处理组间土壤相关农化指标及Fe、Zn元素含量差异无统计学意义,各处理间土壤具有较高的同质性,可保证实验的一致性。
-
如图1所示,在气候变暖与干旱胁迫下,随着胁迫程度的增加,小麦幼苗根、叶部对Fe、Zn元素的富集系数均呈显著的下降趋势,当胁迫程度增加至TD3处理条件时,下降趋势减缓并趋于稳定。在TD1、TD2、TD3处理下,叶部对Fe的富集系数分别下降至对照组的0.73、0.58、0.59倍,对Zn富集系数分别下降至对照组的0.86、0.74、0.75倍;根部对Fe的富集系数分别下降至对照组的0.77、0.59、0.56倍。根部对Zn的富集分别下降至对照组的0.74、0.59、0.60倍。上述结果表明,随着气候变暖和干旱加剧,小麦幼苗对Fe、Zn的富集减少,这意味着在全球变暖背景下,部分地区的Fe、Zn等营养元素缺乏问题或将更加突出。
作物对矿质元素的吸收、富集、迁移和分布受诸多因素的影响和调控,涉及非常复杂的生理生化过程。其中温度与水分作为重要的影响因素已获广泛认可。温度与水分是影响植物生长发育和生理生化代谢的关键因素。研究表明[11-12],干旱与高温复合胁迫能够显著降低卡尔文循环关键光合酶的活性,抑制植物的生长和降低光合速率,也可影响植物细胞的超微结构,改变叶片的气孔性状,减少植物根部的碳分配,同时还可造成质膜过氧化,改变细胞膜的通透性,从而直接或间接影响植物对矿质元素的吸收、分布和利用。温度和干旱也可通过影响土壤矿质元素的可利用性、矿质离子间的相互作用及作物各种生理功能对矿质养分的需求等途径,影响作物养分耦合[13-14] 。李裕等[7]研究发现,3 ℃的增温处理下,马铃薯叶片中Cu、Zn和Fe的浓度分别升高25.0%、27.0% 和24.0%,块茎中、Fe、Zn、Cu的浓度分别下降41.0%、29.0% 和23.0%。增温1℃和2℃的处理下,小麦籽粒中Zn 浓度分别增加了28.9%和35.8%[10]。模拟试验表明,温度升高可降低豌豆和小麦对Fe、Zn的吸收利用[15-16]。由此可见,增温对作物矿质养分的影响尚存在一定的分歧,研究结果因作物种类、元素种类、增温幅度等不同而有所差异,多数研究倾向于增温降低了作物对矿质养分的获取和利用。水分是影响作物获取和利用矿质养分的重要因素,根系对矿质养分的吸收需要呼吸作用的能量驱动,在干旱条件下,植物根系的有氧呼吸会受到显著抑制,进而影响作物对矿质养分的吸收、同化和运输。干旱胁迫也可造成疏水物质在植物根系表皮细胞的胞壁上的积累, 从而影响植物根系对矿质离子吸收和跨膜运输[17]。刘建福等[18]研究表明,干旱胁迫可明显影响澳洲坚果叶片中的矿质元素含量,随胁迫程度增加,叶片中Ca、N、Mg和Zn元素含量显著降低,而K和Mn元素含量则出现升高。干旱胁迫条件下大豆叶片对Fe、Cu累积量均出现降低[14]。李仰锐等[19]研究发现,干旱胁迫下,夏玉米各部位Ca、Mg、Mn元素含量呈现不同程度的增加,Cu元素含量则出现不同程度的降低。显然,关于干旱对作物矿质元素影响的科学认知尚不统一,但多数研究认为,在干旱胁迫下作物对多数矿质元素的获取和利用有所下降,仅少数元素有所增加。目前关于增温与干旱复合胁迫对作物矿质元素的影响研究不多,涉及的元素种类也较少,一些研究发现[20],高温与干旱交互胁迫可显著降低西红柿幼苗叶片、茎的全磷含量,根的全氮含量则显著升高。另有研究表明[21],高温和干旱复合胁迫下刺槐幼苗体内N、P 含量增加,但过度的高温和干旱则会减少P的积累。
结合上述研究分析认为,随着气候变暖与干旱复合胁迫程度的增加,小麦幼苗根、叶部对Fe、Zn元素的富集降低可能原因如下:①增温与干旱复合胁迫降低了小麦幼苗的光合速率和呼吸作用,影响了小麦幼苗对矿质养分的获取和利用能力;②增温与干旱复合胁迫影响了植物体内的水分代谢,由此导致与水分代谢密切相关的矿质元素代谢受到抑制;③增温与干旱复合胁迫降低了土壤的矿质元素可利用性,造成小麦幼苗无法从土壤中有效获取矿质养分。在TD3处理条件下,Fe、Zn元素的富集降低有所减缓并趋于稳定,这可能是由于过高的增温与干旱严重影响了小麦幼苗的生长发育,使得生长“稀释效应”减弱所致,同时也可能和作物的逆境保护机制有关。
-
如图2所示,随着气候变暖与干旱胁迫程度的增加,小麦幼苗根、叶部各亚细胞层Fe、Zn含量相比对照均出现显著的下降,在TD1、TD2、TD3处理下,叶部F1层Fe含量下降幅度为16.45%—31.23%, F2层降幅为33.91%—49.55%, F3层降幅28.84%—42.55%;根部F1层Fe降幅为27.39%—55.67%, F2层降幅为19.70%—32.63%, F3层降幅为 22.83%—53.02%。在TD1、TD2、TD3处理下,叶部F1层Zn含量下降幅度为24.54%—38.17%, F2层降幅为12.94%—25.50%, F3层降幅12.09%—18.02%;根部F1层Zn降幅为27.62%—40.97%, F2层降幅为28.62%—46.56%, F3层降幅为 22.04%—36.68%。在3个亚细胞层中,Fe在小麦根、叶部均以F2层含量较高,Zn在小麦根、叶部均以F3层含量较高,说明Fe在小麦幼苗细胞器中分布较高,Zn在小麦幼苗细胞可溶性组分中分布较高。
上述结果显示,随胁迫程度的增加,小麦幼苗各亚细胞层中的Fe、Zn含量整体上呈现降低趋势,且与根、叶部的整体变化趋势接近,说明增温与干旱胁迫对各亚细胞组织的影响存在一定均衡性。实际上,根、叶部Fe、Zn含量的整体变化在根本上也是决定于其各亚细胞组织对Fe、Zn元素积累的改变。植物细胞对矿质养分的吸收分布需经过根部吸附、质外或共质体吸收、木质部装载、运输和卸载等一系列复杂的生理过程,这一过程同时伴有大量酶及转运蛋白的参与(如NRAMP、FDR3、PIC1、ZIP和ATM等诸多基因家族参与了植物细胞内Fe、Zn的转运[22])。任何外部生境的改变都可能影响和干扰其稳态和平衡,从而影响矿质养分在植物亚细胞中的吸收、分布和功用发挥。Watt等[17]研究表明,干旱胁迫可造成疏水物质在植物根系表皮细胞壁上的积累, 影响矿质元素在质外体中吸收和膜内运输,从而影响各亚细胞组织中矿质元素的动态分布。Gong 等[23]研究认为,干旱导致小麦根系发育受损,活力下降,吸收矿质离子的能力减弱,进而影响小麦对矿质离子的摄取。干旱胁迫也可影响作物体内蛋白质的合成及核酸代谢,降低酶活性,影响植物的矿质养分代谢平衡[20]。温度对植物矿质离子的吸收、转运影响较为复杂,一定范围内的增温有助于提高根部对矿质元素的吸收,同时也有助于促进植物体内相关酶及转运蛋白的合成与活性,但过高的温度则出现相反的效应,且不同植物间的差异较大。结合上述分析认为,增温与干旱复合胁迫可能主要通过影响小麦幼苗的根际环境从而影响矿质元素的根部吸收,同时也可能通过影响与Fe、Zn吸收、运输相关的酶及转运蛋白的合成与活性,进而影响其在亚细胞中的吸收与分布。
-
由图3可见,随着气候变暖与干旱胁迫程度的增加,小麦幼苗根、叶部各亚细胞层Fe、Zn比例分布基本维持在一定范围内波动,仅个别处理组间的差异存在统计学意义。
小麦幼苗叶部F1层Fe分布比例在31.83%—38.82%之间波动;F2层在42.26%—48.89%之间波动;F3层在18.90%—19.33%之间波动。根部F1层Fe在24.73%—31.52%之间波动;F2层在40.77%—50.55%之间波动;F3层在22.19%—27.71%之间波动。根、叶部Fe各亚细胞层分布比例总体排序均为F2 > F1> F3。小麦幼苗叶部F1层Zn分布比例维持在30.68%—36.78%之间;F2层维持在21.62%—23.02%之间;F3层维持在41.62%—47.29%之间。根部F1层Zn维持在在25.34%—26.07%之间;F2层在28.02%—31.12%之间波动;F3层在43.07%—46.24%之间。叶部Zn各亚细胞层分布比例总体排序为F3 > F1 > F2,根部为总体排序为F3 > F2 > F1。
上述研究结果表明,气候变化与干旱胁迫对于小麦幼苗亚细胞中Fe、Zn的比例分布影响具有较高的不确定性,整体上影响有限。这与部分文献报道不同。有研究显示[24],环境温度的改变可以显著影响作物矿质养分在各细胞组分中的分配,温度升高,可明显提高小麦幼苗根部细胞可溶性组分中的Fe、Zn和Mn分配比例,而降低其在细胞壁中的分配比例。Li等[25]研究发现,温度增加也可影响小麦幼苗根部亚细胞组分中Cd的分布。有关干旱对作物矿质元素亚细胞分配的影响研究尚未见公开报道。显然,单一增温胁迫与增温和干旱复合胁迫的效应存在较大差异。一般情况下,多种胁迫因子的组合往往会形成重叠、协同或者拮抗效应。据此分析,温度升高与干旱胁迫两种因素对小麦幼苗矿质元素的亚细胞分布影响可能呈现截然相反的效应,两者的复合胁迫形成拮抗效应,导致其综合效应大为减弱,在结果上表现为各亚细胞层Fe、Zn元素的分布比例维持相对动态稳定。
-
如图4所示,随着气候变暖与干旱胁迫程度的增加,小麦幼苗Fe自根向茎叶部迁移呈现波动趋势,表现为先降低后回升,但整体变化不具有统计学意义。Zn自根向茎叶部的迁移呈现先升高后逐渐稳定的变化趋势。在TD1、TD2和TD3各处理情景下小麦幼苗Fe的转移系数分别为对照组的0.66、0.78、1.06倍,Zn的转移系数分别为对照组的1.15、1.24、1.24倍。
矿质元素在植物体内的迁移和再分布不仅是植物维持正常生长发育和生理代谢平衡的重要手段,同时也是植物响应逆境胁迫的重要反馈和调节机制。吸收进入作物根部的矿质元素,主要依赖木质部导管,随汁液在蒸腾作用的拉力和根压的共同作用下迁移至植物的地上部分,并分配至各个器官,同时也会通过韧皮部进行运输。这一过程的进行也有赖于大量转运蛋白参与。这些转运蛋白在植物矿质元素吸收、木质部装载、卸载及液泡区室化过程中起着极为重要的作用[26]。当植物处在不同逆境条件下时,可能通过激活一系列机制,增强或减弱某些转运蛋白的合成或活性发挥,实现矿质离子的吸收、转运和体内再分配的调控,维持细胞内的离子平衡和体内代谢稳态[27]。温度和水分是影响作物蒸腾作用发挥的关键因素,研究表明[28],干旱可造成植物的蒸腾作用降低,从而影响矿质元素向地上部的迁移。而一定范围内的增温则有助于提高作物的蒸腾作用,促进植物体内矿质养分的向上运输[29]。据此来看,增温与干旱复合胁迫对小麦幼苗蒸腾作用的影响尚具有一定的不确定性。本研究结果显示,小麦幼苗Fe、Zn自根向叶部迁移呈现不同的变化,显然,这一现象并不能简单的归因于蒸腾作用的改变使然,更为可能是增温和干旱复合胁迫在不同程度上影响了Fe、Zn相关转运蛋白的合成及活性发挥,从而表现为Fe、Zn自根部向地上部迁移的改变,由于参与Fe、Zn转运的蛋白种类不同,其迁移变化趋势则表现不同。
-
随着温度增加和干旱的加剧,小麦幼苗根、叶部对Fe、Zn的富集均呈明显的减少趋势,当胁迫程度达到较高的程度时,下降趋势有所减缓并趋于稳定;小麦幼苗根、叶部各亚细胞组分中的Fe、Zn的含量随温度和干旱复合胁迫水平的提高均出现不同程度的下降;Fe在小麦幼苗细胞器中分布比例较高,Zn在小麦幼苗细胞可溶性组分中分布比例较高;增温与干旱复合胁迫对小麦幼苗根、叶部各亚细胞组分中Fe、Zn分布比例影响有限,显著性不高;随着胁迫水平的增加,小麦幼苗Fe自根向茎叶部迁移呈现波动趋势, 表现为先降低后回升, Zn自根向茎叶部迁移呈现先升高后逐渐稳定的变化趋势。
模拟气候变暖与干旱复合胁迫对小麦幼苗亚细胞水平Fe、Zn动态积累的影响
Effects of simulated climate warming and drought combined stress on dynamic accumulation of Fe and Zn at subcellular level in wheat seedlings
-
摘要: 为了解气候变化背景下,作物矿质养分利用在亚细胞水平的响应动态及其机理,本研究在人工气候室通过模拟实验,开展了气候变暖与干旱复合胁迫对小麦幼苗亚细胞水平Fe、Zn动态积累影响的探索. 研究显示,随着胁迫程度增加,小麦幼苗根、叶部对Fe、Zn元素的富集均呈显著下降趋势,根、叶部Fe最低分别下降至对照的0.56倍和0.58倍,Zn最低分别下降至对照的0.59倍和0.74倍. 小麦幼苗根、叶部各亚细胞组分中的Fe、Zn含量随胁迫水平的提高均呈现不同程度下降,各亚细胞分布比例变化不明显. Fe自根向茎叶部迁移呈现先降低后回升的趋势,各处理Fe转移系数分别为对照组的0.66、0.78、1.06倍;Zn自根向茎叶部的迁移呈现先升高后逐渐稳定的趋势,各处理Zn转移系数分别为对照组的1.15、1.24、1.24倍. 结果表明, 增温与干旱复合胁迫可在不同层面对小麦幼苗Fe、Zn元素的动态富集产生影响,最终可能对农业生产及人类健康产生不利影响. 本研究将为农业系统科学应对气候变化提供参考。Abstract: In order to understand the dynamics response and mechanism of mineral nutrient utilization of crop at the subcellular level under the background of climate change, a simulated experiment was conducted in the artificial climate chamber to evaluate the combined effects of climate warming and drought stress on the bio-accumulation of Fe and Zn at subcellular level in Wheat Seedlings. The results showed with the increase of stress degree, the enrichment of Fe and Zn in roots and leaves of wheat seedlings had a significant downward trend. The lowest content of Fe in roots and leaves decreased to 0.56 and 0.58 times of the control group, Zn decreased to 0.59 and 0.74 times of the control group, respectively. The contents of Fe and Zn in the subcellular components of roots and leaves of wheat seedlings decreased with the increase of stress level. However, the proportion of subcellular distribution showed no significant change. The migration of Fe from root to stem and leaf showed a trend of first decreasing and then rising, the transfer coefficients of Fe in each treatment were 0.66, 0.78 and 1.06 times that of the control group, respectively. The migration of Zn from root to stem and leaf showed a trend of first increasing and then gradually stabilizing, the transfer coefficients of Zn in each treatment were 1.15, 1.24 and 1.24 times that of the control group, respectively. The results showed that the combined stress of warming and drought could affect the dynamic enrichment of Fe and Zn in wheat seedlings at different levels, which would eventually have an adverse impact on agricultural production and human health. This study will provide a reference for the agricultural system to deal with climate change scientifically.
-
Key words:
- climate warming /
- drought /
- wheat seedlings /
- Fe and Zn
-

-
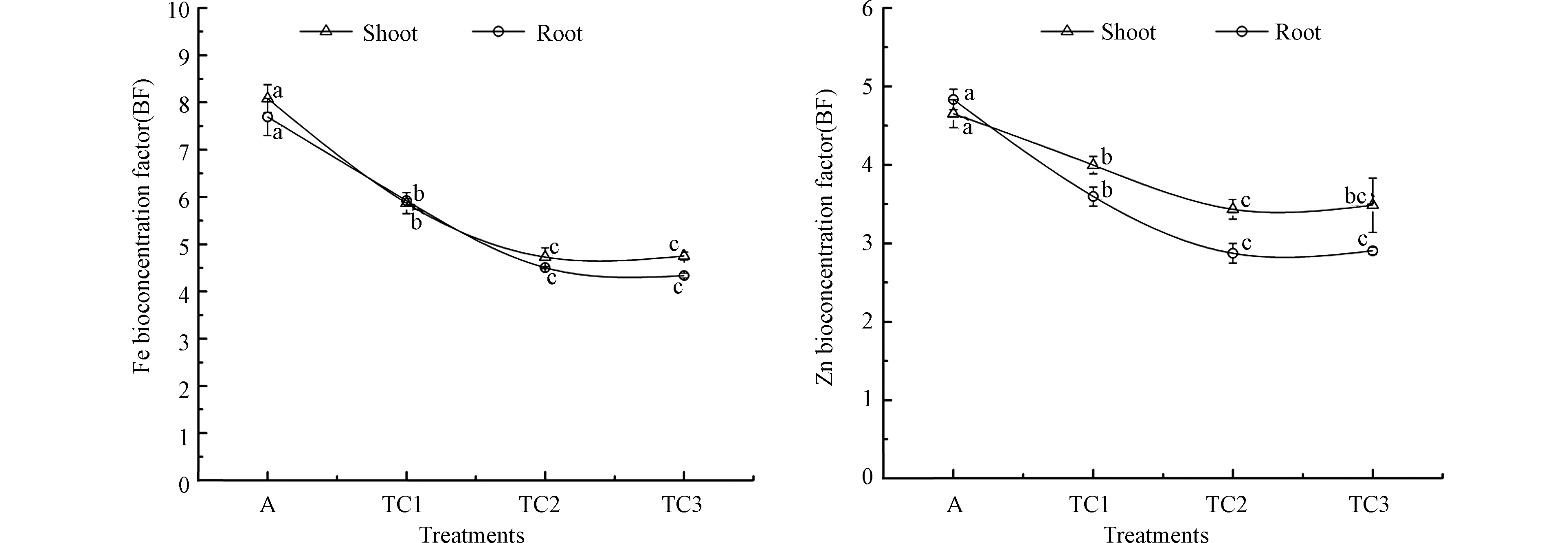
图 1 气候变暖与干旱胁迫对小麦幼苗根、叶部Fe、Zn富集的影响
Figure 1. The combined effects of warming and drought on the BF values of Fe and Zn in wheat shoots and roots
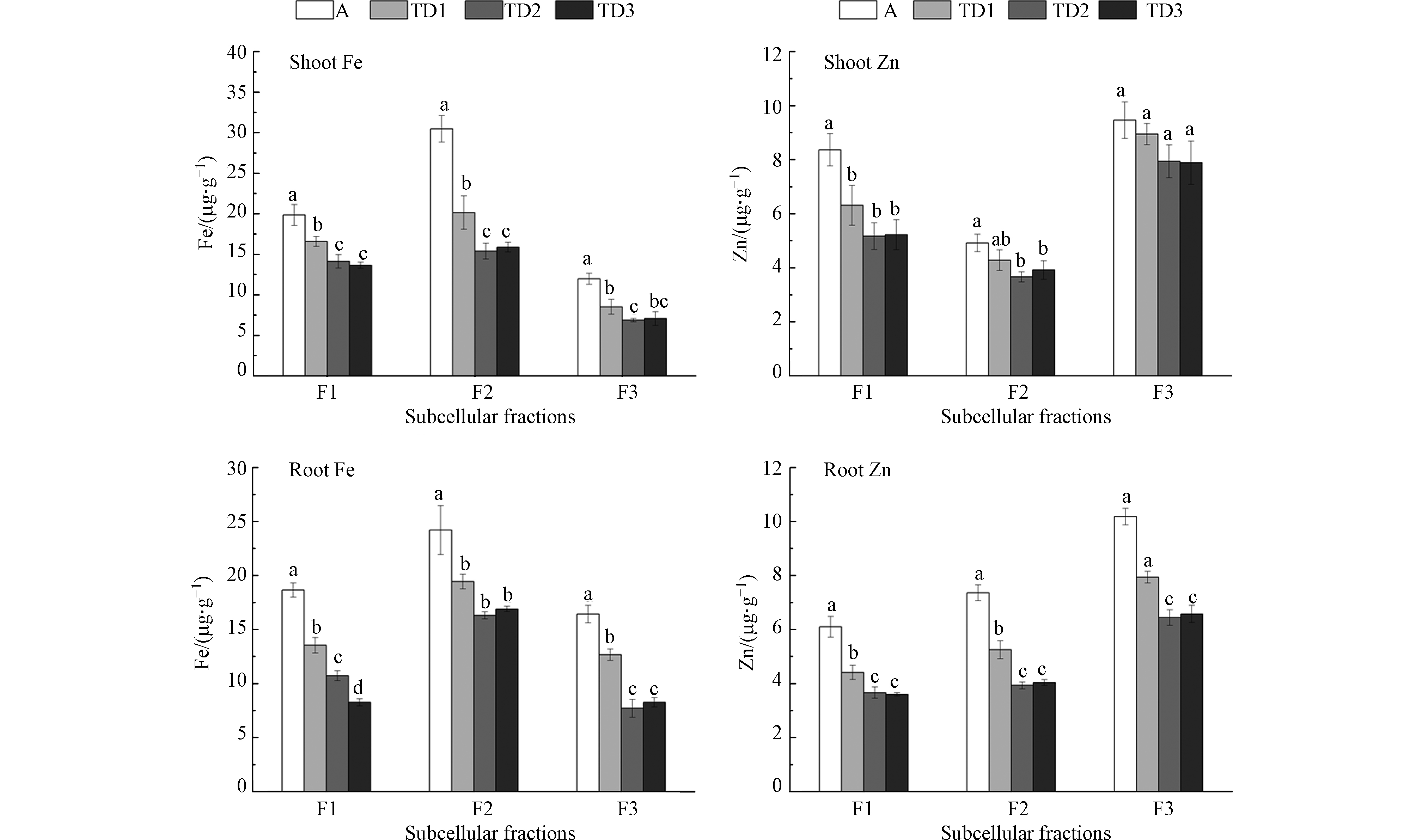
图 2 气候变暖与干旱胁迫对小麦幼苗亚细胞Fe、Zn含量的影响
Figure 2. The combined effects of warming and drought on the subcellular concentrations of Fe and Zn in wheat shoots and roots
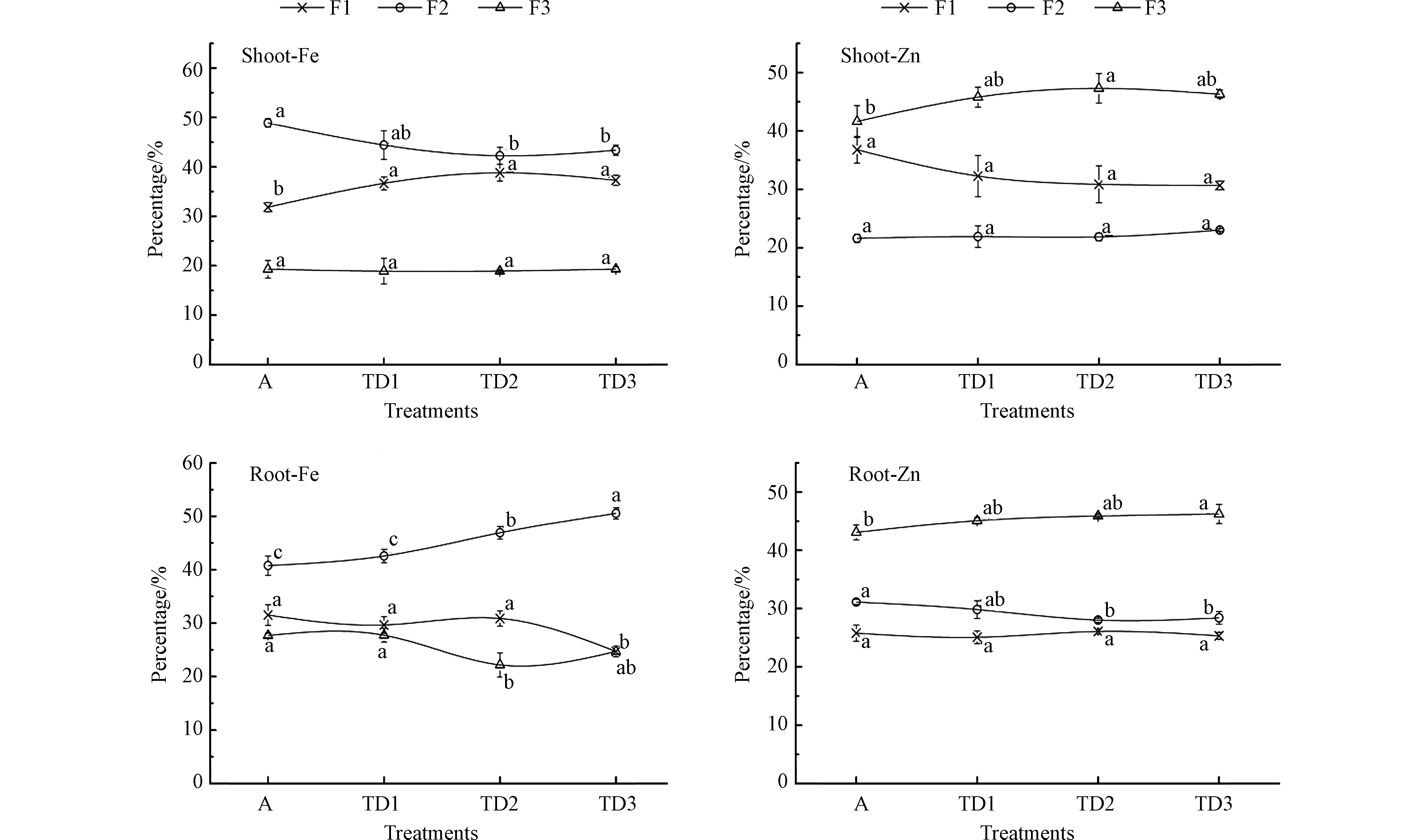
图 3 气候变暖与干旱胁迫对小麦幼苗根、叶部Fe、Zn亚细胞分布比例的影响
Figure 3. The combined effects of warming and drought on the proportional distribution of Fe and Zn in the subcellular fractions of wheat roots and shoots
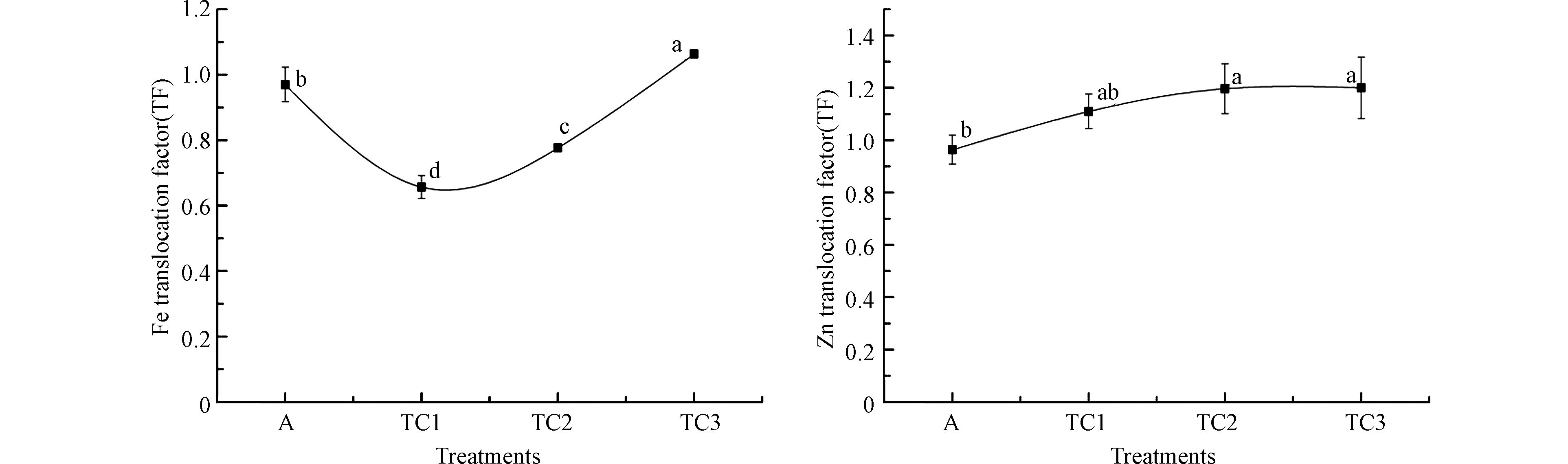
图 4 气候变暖与干旱胁迫对小麦幼苗Fe、Zn转移系数的影响
Figure 4. The combined effects of warming and drought on the translocation factor ( TF)
表 1 模拟处理条件和对照设计详表
Table 1. Details about the three simulated climate treatments and the untreated control condition
A TD1 TD2 TD3 气温升高/℃
Temperature rise value+0 +1 +2 +3 土壤持水量
Relative soil moisture capacity70%—75% 60%—65% 45%—50% 30%—35%  下载: 导出CSV
下载: 导出CSV
表 2 实验前供试地耕作层(0—20 cm)土壤农化性质
Table 2. Soil agrochemical properties of the cultivated layer (0—20 cm) of the test land before the experiment
处理
Treatments有机质/(g·kg−1)
Organic
matter总氮/(g·kg−1)
Total
nitrogen碱解氮/(mg·kg−1)
Alkali-hydro
nitrogen速效磷/(mg·kg−1)
Rapidly-available
phosphorus速效钾/(mg·kg−1)
Rapidly-available
potassiumpH值
pHA 10.60±0.68 a 0.89±0.11 a 79.54±7.76 a 28.52±2.01 a 296.39±17.46 a 7.96±0.19 a TD1 10.54±0.91 a 0.82±0.09 a 83.16±6.41 a 29.65±1.91 a 293.58±13.68 a 8.04±0.29 a TD2 10.36±0.92 a 0.83±0.09 a 86.11±7.66 a 29.82±2.24 a 296.57±13.50 a 7.97±0.26 a TD3 10.48±0.75 a 0.89±0.15 a 82.52±8.53 a 30.42±2.41 a 292.64±12.60 a 7.89±0.31 a 注:表中数据为5个样点的平均值±标准差,各列4种处理间相同字母代表无统计学显著性差异,pH值测定在水土比5︰1 条件下进行.
Note: the data in the table are the average value ± standard deviation of five sample points; the same letter between the four treatments in each column represents no statistically significant difference. The pH value was measured under the condition of water soil ratio of 5:1.
下载: 导出CSV
表 3 供试地耕作层(0—20cm)土壤元素有效态含量
Table 3. Available state content of soil elements in the cultivated layer (0—20cm) of the test land
处理Treatments A TD1 TD2 TD3 Fe/(μg·g−1) 7.71±1.07a 6.92±0.62 a 6.87±0.77 a 7.63±0.64 a Zn/(μg·g−1) 4.89±0.34 a 5.26±0.54 a 5.35±0.40 a 5.22±0.67a 注:表中数据为5个样点的平均值±标准差,各行4种处理间相同字母代表无统计学显著性差异.
Note: the data in the table are the average value ± standard deviation of five sample points; the same letter between the four treatments in each column represents no statistically significant difference.
下载: 导出CSV
-
[1] 中国气象局气候变化中心. 中国气候变化蓝皮书2021 [M]. 北京: 科学出版社, 2021. Climate Change Center of China Meteorological administration. Blue book on climate change in China 2021 [M]. Beijing: Science Press, 2021(in Chinese).
[2] 王馥棠. 气候变化对农业生态的影响 [M]. 北京: 气象出版社, 2003. WANG F T. Impact of climate change on agricultural ecology [M]. Beijing: China Meteorological Press, 2003(in Chinese).
[3] DAI A G. Drought under global warming: A review [J]. Wiley Interdisciplinary Reviews:Climate Change, 2011, 2(1): 45-65. doi: 10.1002/wcc.81 [4] 方霞. 中国干旱半干旱生态系统生产力和碳动态对气候变化响应的数值模拟研究 [D]. 乌鲁木齐: 新疆大学, 2018. FANG X. A numeric study on the impact of climate change on ecosystem carbon dynamics and productivity in China's arid and semi-arid regions [D]. Urumqi: Xinjiang University, 2018(in Chinese).
[5] BEACH R H, SULSER T B, CRIMMINS A, et al. Combining the effects of increased atmospheric carbon dioxide on protein, iron, and zinc availability and projected climate change on global diets: A modelling study [J]. The Lancet Planetary Health, 2019, 3(7): e307-e317. doi: 10.1016/S2542-5196(19)30094-4 [6] 吕亮杰, 刘少兴, 陈希勇, 等. 干旱胁迫下不同类型小麦灌浆期营养物质积累动态 [J]. 华北农学报, 2021, 36(2): 89-98. doi: 10.7668/hbnxb.20191708 LÜ L J, LIU S X, CHEN X Y, et al. Nutrient accumulation dynamics of different types of wheat during grain filling stage under drought stress [J]. Acta Agriculturae Boreali-Sinica, 2021, 36(2): 89-98(in Chinese). doi: 10.7668/hbnxb.20191708
[7] LI Y, ZHANG Q, WANG R Y, et al. Temperature changes the dynamics of trace element accumulation in Solanum tuberosum L [J]. Climatic Change, 2012, 112(3/4): 655-672. [8] SARDANS J, PEÑUELAS J, ESTIARTE M. Warming and drought change trace element bioaccumulation patterns in a Mediterranean shrubland [J]. Chemosphere, 2008, 70(5): 874-885. doi: 10.1016/j.chemosphere.2007.06.085 [9] WANG X H, LI Y, LU H, et al. Combined effects of elevated temperature and CO2 concentration on Cd and Zn accumulation dynamics in Triticum aestivum L [J]. Journal of Environmental Sciences, 2016, 47: 109-119. doi: 10.1016/j.jes.2015.11.029 [10] 李裕, 张强, 王润元, 等. 气候变暖对春小麦籽粒痕量元素利用率的影响 [J]. 农业工程学报, 2011, 27(12): 96-104. doi: 10.3969/j.issn.1002-6819.2011.12.019 LI Y, ZHANG Q, WANG R Y, et al. Influence of climatic warming on accumulation of trace elements in spring wheat (Triticum aestivum L. ) [J]. Transactions of the Chinese Society of Agricultural Engineering, 2011, 27(12): 96-104(in Chinese). doi: 10.3969/j.issn.1002-6819.2011.12.019
[11] 肖万欣, 王延波, 谢甫绨, 等. 高温和干旱胁迫下, 植物叶片光合系统变化、气孔运动及其调控研究进展 [J]. 辽宁农业科学, 2012(6): 29-33. doi: 10.3969/j.issn.1002-1728.2012.06.008 XIAO W X, WANG Y B, XIE F T, et al. Research progress of photosystem variation, stoma movement and its regulation of plant leaf under high temperature and drought stress [J]. Liaoning Agricultural Sciences, 2012(6): 29-33(in Chinese). doi: 10.3969/j.issn.1002-1728.2012.06.008
[12] 王雅晴. 不同小麦品种对高温干旱及共胁迫的响应机制 [D]. 泰安: 山东农业大学, 2019. WANG Y Q. Response mechanism of different wheat cultivars to high temperature, drought and their combined stress [D]. Taian: Shandong Agricultural University, 2019(in Chinese).
[13] 李裕, 王小恒, 罗兴平, 等. 气候变化对半干旱地区作物微量元素利用率影响研究进展 [J]. 干旱气象, 2020, 38(6): 895-899,1030. LI Y, WANG X H, LUO X P, et al. Influence of climate change on bioavailability of micronutrients in crops in semiarid area [J]. Journal of Arid Meteorology, 2020, 38(6): 895-899,1030(in Chinese).
[14] 赵立琴, 龚振平, 马春梅. 干旱胁迫对大豆叶片矿质元素含量和累积量影响 [J]. 东北农业大学学报, 2013, 44(2): 12-16. doi: 10.3969/j.issn.1005-9369.2013.02.005 ZHAO L Q, GONG Z P, MA C M. Effect of drought stress on concentration and accumulation of mineral nutrition in leaves of soybean seedlings [J]. Journal of Northeast Agricultural University, 2013, 44(2): 12-16(in Chinese). doi: 10.3969/j.issn.1005-9369.2013.02.005
[15] KUMARI M, VERMA S C, BHARDWAJ S K. Impact of elevated CO2 and temperature on quality and biochemical parameters of pea (Pisum sativum) [J]. Indian Journal of Agricultural Science, 2017, 87(8): 1035-1040. [16] HOUSHMANDFAR A, FITZGERALD G J, MACABUHAY A A, et al. Trade-offs between water-use related traits, yield components and mineral nutrition of wheat under Free-Air CO2 Enrichment (FACE) [J]. European Journal of Agronomy, 2016, 76: 66-74. doi: 10.1016/j.eja.2016.01.018 [17] WATT M, WEELE C M, MCCULLY M E, et al. Effects of local variations in soil moisture on hydrophobic deposits and dye diffusion in corn roots [J]. Botanica Acta, 1996, 109(6): 492-501. doi: 10.1111/j.1438-8677.1996.tb00602.x [18] 刘建福, 倪书邦, 贺熙勇, 等. 水分胁迫对澳洲坚果叶片矿质元素含量的影响 [J]. 热带农业科技, 2004, 27(1): 1-3. doi: 10.3969/j.issn.1672-450X.2004.01.001 LIU J F, NI S B, HE X Y, et al. Effect of moisture stress on mineral content in the Macadamia leaf [J]. Tropical Agricultural Science & Technology, 2004, 27(1): 1-3(in Chinese). doi: 10.3969/j.issn.1672-450X.2004.01.001
[19] 李仰锐, 隋方功, 徐卫红, 等. 水分胁迫对夏玉米(收获期)中微量元素含量的影响 [J]. 莱阳农学院学报, 2005, 22(2): 115-117. LI Y R, SUI F G, XU W H, et al. Effects of water stress on contents of mid-element and micro-element in summer maize (harvest stage) [J]. Journal of Laiyang Agricultural College, 2005, 22(2): 115-117(in Chinese).
[20] 王德福, 段洪浪, 黄国敏, 等. 高温和干旱胁迫对西红柿幼苗生长、养分含量及元素利用效率的影响 [J]. 生态学报, 2019, 39(9): 3199-3209. WANG D F, DUAN H L, HUANG G M, et al. Effects of high temperature and drought stress on growth, nutrient concentration, and nutrient use efficiency of tomato seedlings [J]. Acta Ecologica Sinica, 2019, 39(9): 3199-3209(in Chinese).
[21] 邵维, 吴永波, 杨静, 等. 高温干旱复合胁迫对刺槐幼苗生理生化性能的影响 [J]. 中国农学通报, 2014, 30(4): 1-7. doi: 10.11924/j.issn.1000-6850.2013-2438 SHAO W, WU Y B, YANG J, et al. The combined stress of elevated temperature and drought on the physiological and biochemical characteristics of Robinia pseudoacacia L. seedlings [J]. Chinese Agricultural Science Bulletin, 2014, 30(4): 1-7(in Chinese). doi: 10.11924/j.issn.1000-6850.2013-2438
[22] 虞银江, 廖海兵, 陈文荣, 等. 水稻吸收、运输锌及其籽粒富集锌的机制 [J]. 中国水稻科学, 2012, 26(3): 365-372. YU Y J, LIAO H B, CHEN W R, et al. Mechanism of Zn uptake, translocation in rice plant and Zn-enrichment in rice grain [J]. Chinese Journal of Rice Science, 2012, 26(3): 365-372(in Chinese).
[23] GONG J M, LEE D A, SCHROEDER J I. Long-distance root-to-shoot transport of phytochelatins and cadmium in Arabidopsis [J]. Proceedings of the National Academy of Sciences of the United States of America, 2003, 100(17): 10118-10123. doi: 10.1073/pnas.1734072100 [24] 翁南燕, 周东美, 武敬, 等. 铜镉复合胁迫下温度对小麦幼苗生长及其对铜、镉和矿质营养元素吸收与各元素在亚细胞分布的影响 [J]. 生态毒理学报, 2011, 6(6): 607-616. WENG N Y, ZHOU D M, WU J, et al. Uptake, subcellular distributions of Cu, Cd and mineral elements, and plant growth for wheat seedlings under stress of Cu and Cd as affected by temperature [J]. Asian Journal of Ecotoxicology, 2011, 6(6): 607-616(in Chinese).
[25] LI D D, ZHOU D M, WANG P, et al. Temperature affects cadmium-induced phytotoxicity involved in subcellular cadmium distribution and oxidative stress in wheat roots [J]. Ecotoxicology and Environmental Safety, 2011, 74(7): 2029-2035. doi: 10.1016/j.ecoenv.2011.06.004 [26] 孙瑞莲, 周启星. 高等植物重金属耐性与超积累特性及其分子机理研究 [J]. 植物生态学报, 2005, 29(3): 497-504. doi: 10.3321/j.issn:1005-264X.2005.03.022 SUN R L, ZHOU Q X. Heavy metal tolerance and hyperaccumulation of higher plants and their molecular mechanisms: A review [J]. Acta Phytoecologica Sinica, 2005, 29(3): 497-504(in Chinese). doi: 10.3321/j.issn:1005-264X.2005.03.022
[27] 余丹萍, 李取生, 王立立, 等. 缺Fe/Zn及盐胁迫下苋菜对Cd及矿质元素的吸收与IRT1表达的关系 [J]. 农业环境科学学报, 2016, 35(2): 234-239. doi: 10.11654/jaes.2016.02.004 YU D P, LI Q S, WANG L L, et al. Cadmium and mineral element accumulation and IRT1 gene expression of Amaranth hybidus L. under iron/zinc deficiencies or salt stress [J]. Journal of Agro-Environment Science, 2016, 35(2): 234-239(in Chinese). doi: 10.11654/jaes.2016.02.004
[28] 白艳波, 李娇, 张宝龙, 等. 干旱胁迫对植物矿质元素影响的研究进展 [J]. 生物技术通报, 2013(3): 15-18. BAI Y B, LI J, ZHANG B L, et al. Research advance on effect of drought stress on mineral elements of plant [J]. Biotechnology Bulletin, 2013(3): 15-18(in Chinese).
[29] 屠小菊, 汪启明, 饶力群. 高温胁迫对植物生理生化的影响 [J]. 湖南农业科学, 2013(13): 28-30. TU X J, WANG Q M, RAO L Q. Effects of high temperature stress on physiology and biochemistry of plant [J]. Hunan Agricultural Sciences, 2013(13): 28-30(in Chinese).
-
 点击查看大图
点击查看大图
计量
- 文章访问数: 2282
- HTML全文浏览数: 2282
- PDF下载数: 59
- 施引文献: 0
