


-
在我国常见的慢性肾脏病中,糖尿病肾病(diabetic nephropathy,DN)位居第一,有研究报告指出,由DN导致的尿毒症高峰或将在20年内来临[1]. 糖尿病作为以高血糖为主要特点的慢性代谢性疾病,机体异常血糖环境可通过炎症反应、氧化应激、促凝血以及内皮功能障碍等引起一系列并发症[2],而DN作为糖尿病的严重微血管并发症之一[3],因其发病机制复杂、特异性药物较少和早期诊断缺乏,给我国带来了严重的经济负担和社会压力[4],据流行病学调查显示,随着我国糖尿病患病人数的增加和患病人群逐渐低龄化,约30%—40%的糖尿病患者合并DN[5]. 并且与无并发症糖尿病患者的平均222美元的治疗费用相比,DN患者的平均治疗费用高达603美元[6],DN已经成为值得关注的公共卫生问题. 相关研究表明,DN的发生发展是遗传因素和环境因素相互作用的结果[7],因此,从环境病因学入手探讨环境污染物暴露与DN的关联对DN发病机制研究和不同人群精准预防十分必要.
环境污染物是指进入环境后使环境的正常组成和性质发生直接或间接变化并对人体健康有害的物质,包括重金属、持久性有机污染物、农药和微塑料等一系列污染物[8]. 随着世界城市化进程的加快,日常生活环境中接触的空气、食物、水中均有污染物的存在,无疑增加了人们与环境暴露污染物的接触率. 环境污染物可通过各种方式影响生物进程,并损伤人体健康. Huang等[9]研究表示,随着机体内血清铅浓度的上升,其肾小球滤过率逐渐下降,揭示铅负荷对Ⅱ型糖尿病患者的进行性DN有长期影响. 一项嵌套病例对照研究显示[10],有机污染物与糖尿病具有相关性,但与其发生率并无明显关系,并且还指出多氯联苯等会增加DN死亡风险. Everett等[11]评估了1999—2004年全国健康和营养检查调查中血液中6种氯化二苯并二噁英、9种氯化二苯并呋喃和8种多氯联苯与DN的相关性,结果显示暴露于二噁英、呋喃和多氯联苯会导致DN的发生和(或)恶化. 虽然这些研究证明了环境污染物与DN发生发展甚至DN患者死亡率之间都存在关系,但是都通过DN的病理生理状态改变如肾小球滤过率下降、尿白蛋白水平增高和肾组织纤维化等最终指标进行评价,对于更微观的环境污染物如何干预DN的分子机制并没有解释[12]. 另外,现有的实验研究还存在样本缺乏[13]、环境污染物单一、多种环境污染物研究无法确认主次之分[10]和无法排除混杂因素[14]如基因变异、其他污染物影响以及药物使用等问题,而从基因层面入手对环境污染物进行分析,可以降低样本缺乏对结果的影响,以及解决研究环境污染物单一和无法确定多种环境污染物研究主次之分的缺陷,同时也不用担忧无法排除的混杂因素对结果的影响,从而更好的探明环境污染物与DN发生发展的关系.
生物信息学作为一门新兴交叉学科,其衍生出的包括基因组学、转录组学、蛋白组学等在内的一系列组学研究贯穿于疾病发生机制、药物研发、分子诊断以及疾病预防领域[15]. Van等[16]则通过蛋白质组学结合生物信息学探究了可以预测DN不同发展阶段的生物标志物,还有研究[17]则在生物信息学的基础上联合meta分析,确定了DN的易感基因及其富集的信号通路. 随后,环境污染物相关的生物信息学逐渐兴起. Verga等[18]通过生物信息学方法全基因组测序并分析了暴露于内分泌干扰物对动物及人的不良影响和分子途径. 还有研究[19]通过生物信息学方法构建了包含27个人体组织中检测到的380种环境污染物的人体组织特异性暴露体图谱,并进一步研究了污染物暴露与疾病的关系,揭示了疾病合并症的可能. 以上既往研究都表明了生物信息学方法用于研究多种环境污染物与疾病的相互关系及其分子作用机制的可行性,从而为通过生物信息学方法研究环境污染物暴露与DN的关联性提供了一定参考. 但是,以往研究并没有对多种环境污染物暴露影响疾病的轻重程度进行主次之分,得到的都是基于多种环境污染物的统一结果,并没有进行分类和下一步处理,这将无法区分哪种或者哪几种环境污染物对某疾病影响最严重,造成精准预防的失措. 另外,还有研究提出了通过生物信息学预测生物标志物以及在研究疾病环境病因学时提出环境污染物暴露引起原发疾病合并症的可能,但对于环境污染物暴露是否真的在原发疾病基础上诱发合并症的产生并没有深入的研究,甚至是原发病的生物标志物能否用于合并症的评估也不曾说明. 那么,如何在对不同来源的大数据样本进行挖掘获取其共同特征的同时,构建出需优先回避或干预的环境污染物清单及将其与DN和合并症联合起来共同研究是非常必要的.
故此,本研究将利用生物信息学方法在挖掘影响DN的全氟辛烷磺酸盐、双酚类、有机磷农药、有机氯农药等主要环境污染物的同时还对环境污染物影响DN的主要基因进行评估,预测其表达水平对DN合并肾癌的预后关系,这对环境污染物暴露影响DN研究及预防和早期筛查DN合并肾癌具有一定意义.
-
分别以检索词“(Diabetic nephropathy)AND "Homo sapiens" [porgn:_txid9606]”、“Diabetic nephropathy”在GEO(https://www.ncbi.nlm.nih.gov/geo/)和CTD(http:∥ctdbase.org/)数据库进行检索. 在GEO数据库中,选取无药物等其他条件干预、除DN外无其他合并疾病的含有DN和非DN患者样本的基因芯片数据集纳入本次研究(GSE1009、GSE142153、GSE4117、GSE47183、GSE47184、GSE30528、GSE30529、GSE46899和GSE46897),先通过GEO2R附带箱形图判断选取样本数据是否标准化,如箱形图的值以中位数为中心则表示所选数据已标准化且可以交叉比较,如果显示未标准化,则通过在GEO附带的Options中选择Force normalization选项进行强制规范化,从而使选中的对比样本具有相同的值分布. 对所选样本进行标准化处理后,导出GEO2R分析结果,以adj P<0.05且|log Fold change|>1为筛选条件获取DN和非DN患者的差异表达基因. 在CTD数据库中,下载影响DN进展的基因和环境污染物.
-
利用Venny2.1在线软件(https://bioinfogp.cnb.csic.es/tools/venny/)将GEO和CTD所得基因取交集即获得环境污染物影响DN的共同关联基因,将其导入DAVID数据库(https://david.ncifcrf.gov/)进行生物富集分析,将GO富集分析中生物学过程(biological process,BP)、细胞组成(celluar component,CC)和分子功能(molecular function,MF)3个部分各取Pvalue最小的前10条进行可视化,KEGG富集则选择Pvalue最小的前20条进行可视化,探讨环境污染物影响DN进展的分子机制.
-
利用String数据库(https://cn.string-db.org/)对环境污染物影响DN的基因进行蛋白互作分析,设置物种为“Homo sapiens”,置信度得分阈值设置为0.4,以TSV格式文件导出后再导入Cytoscape3.8.2软件,利用其插件CytoHubba中的最大集团中心度(maximal clique centrality,MCC)算法筛选出Top200基因作为关键基因. MCC算法是已被证实的CytoHubba插件11中拓扑分析方法中预测关键靶点的较为准确的方法,通过该算法可得到环境污染物影响DN的关键靶点并有利于后续的环境污染物分析. 对Top200关键基因中的前20个核心基因进行PPI可视化.
-
将Top200基因导入Metascape在线分析平台(http:∥metascape.org/),通过Mcode模块功能分析关键基因聚集的板块及影响各板块的环境污染物. 通过计算各个模块基因对应的环境污染物频次进行排序,出现次数越多的越靠前,代表该环境污染物影响的靶点基因个数越多,即在影响DN进程的环境污染物中越重要,最后再将各板块的环境污染物进行汇总,选出影响DN进程最主要的环境污染物,体现环境污染物影响DN发生发展主次之分的同时构建出可供参考的优先回避和干预环境污染物清单.
-
为了进一步验证主要环境污染物对关键基因的直接影响及作用位点,将Top20的关键基因蛋白作为小分子配体,主要环境污染物为大分子受体,从PDB数据库(
http://www.rcsb.org/ )下载关键基因的3D结构,通过Pubchem数据库(http://pubchem.ncbi.nlm.nih.gov/ )获取主要环境污染物的pdb格式,通过Open Babel软件将其转化成Mol2格式. 利用Autodock Tools对关键蛋白进行脱水、加氢预处理后进行分子对接. 当结合能<−4.5 kcal·mol−1时代表受体与配体结合活性良好,最后将Autodock Tools获得的pdbqt结果文件导入Open Babel软件转化为pdb格式,利用Discovery Studio Visualizer软件对结合能最高的关键蛋白进行可视化,阐明主要环境污染物与关键基因蛋白的主要结合位点和作用力. -
对环境污染物影响DN的Top20关键基因在肾透明细胞癌(Kidney renal clear cell carcinoma,KIRC)组织中的表达水平进行分析,进而研究环境污染物是否会在影响DN进程时进一步诱发肾癌. 将Top20关键基因分别导入GEPIA在线分析平台(http://gepia.cancer-pku.cn/),在GEPIA2的Expression Analysis部分选择Expression DIY,用箱形图进行展示,癌症类型选择为KIRC,其他默认原始设置,进行表达分析.
-
在基因表达分析的基础上进一步研究Top20关键基因与KIRC患者预后的关系,有利于DN合并KIRC早期诊断和预后推断的同时对精确靶向治疗也具有重要意义. 利用Kaplan-Meier Plotter(
http://www.kmplot.com/ )对Top20关键基因进行生存分析,生存率的比较采用Log-rank检验,设置Auto select best cutoff区分高表达组和低表达组,总生存期(overall survival,OS)绘制KM生存曲线,选择癌症类型为KIRC,其他默认原始设置,P<0.05即具有统计学意义. -
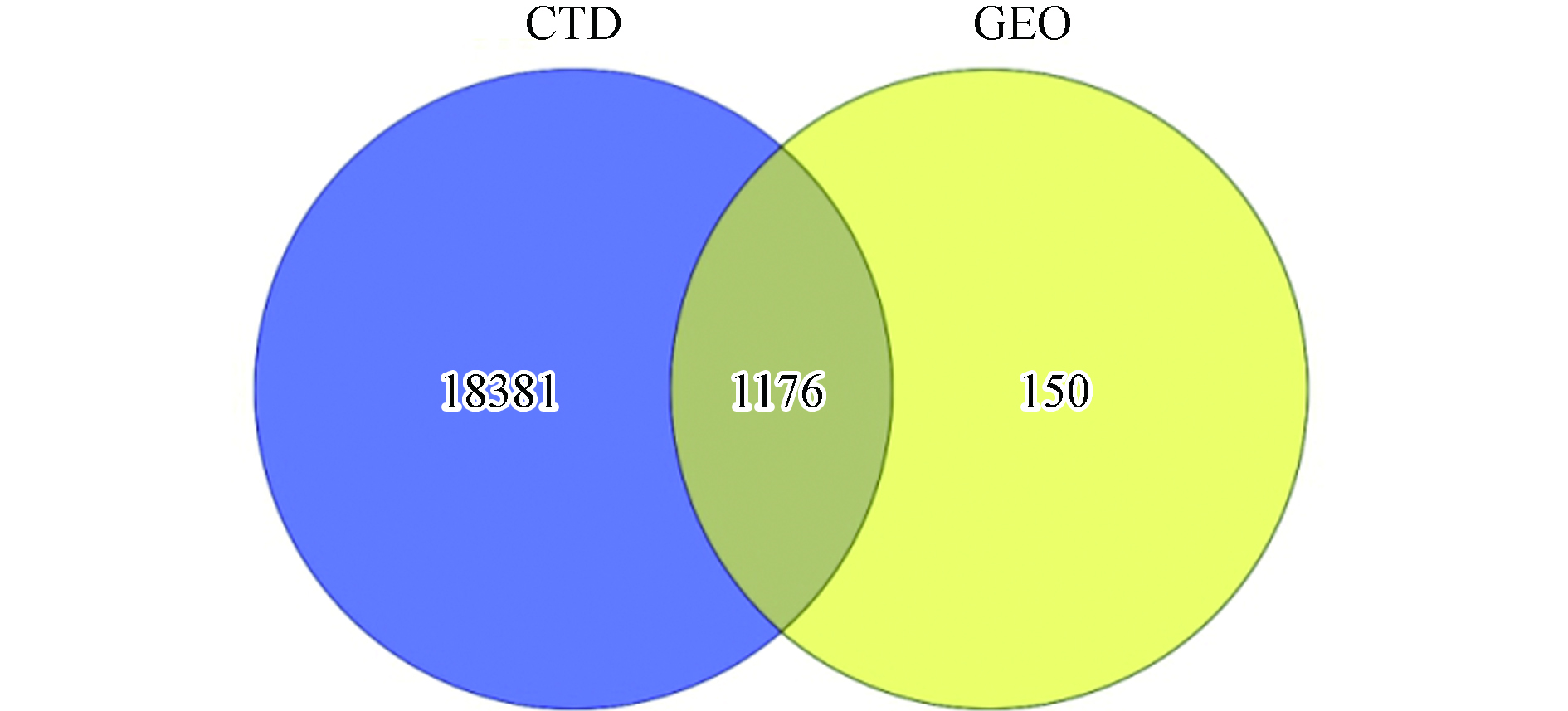
经过GEO数据库检索筛选,共获得9组与DN相关联的基因芯片数据集,其芯片数据详细情况见表1,共涉及345个样本,其中取材主要为肾组织活检和外周血,并且大部分数据集来自美国,纳入男性例数略高于女性. 其中,数据集GSE47185、GSE30122和GSE46900都有对应的子数据集,后续研究都通过子数据集进行分析,另外GPL14663是数据集GSE47185的分析平台之一,用于本次研究的数据都以子数据集GSE47183和GSE47184于GPL14663平台分析结果为基础. 确定研究数据集后经过标准化处理,进行差异基因筛选,再结合CTD数据库检索结果,两者取交集得到1176个环境化学物影响DN的共同关联基因(图1).
-
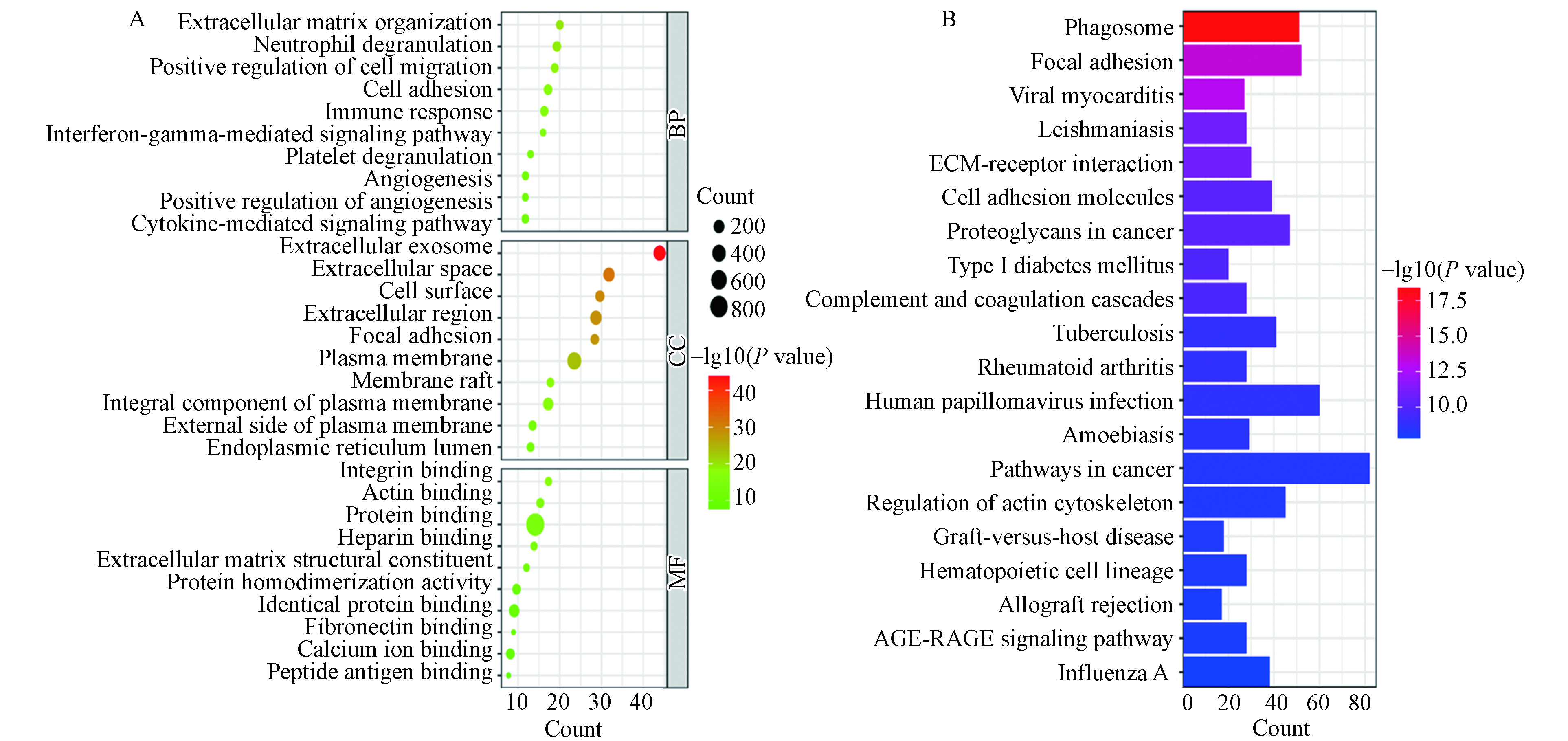
为了明确环境污染物影响DN基因所涉及的生物学过程及富集通路,对1176个共同关联基因进行生物富集分析. 将BP、CC、MF中Pvalue最小的前10条进行GO功能富集可视化,如图2A所示,圆点越大则代表该生物功能富集的共同关联基因就越多,颜色越接近红色则代表Pvalue越小. 而在GO功能富集分析中,以BP最为重要,由图2A可知,环境污染物影响DN的基因主要富集于中性粒细胞脱颗粒(neutrophil degranulation)、细胞迁移正调控(positive regulation of cell migration)、细胞黏附(cell adhesion)、免疫反应(immune response)、血小板脱颗粒(platelet degranulation)及血管生成正调控(positive regulation of angiogenesis)等生物过程,进而提示环境污染物可通过炎症反应和影响机体组织血管生成加速DN进展.
将KEGG信号通路富集结果中Pvalue最小的前20条进行可视化,如图2B所示,条形越长代表该信号通路富集的共同关联基因就越多,颜色越接近红色代表Pvalue越小. 由图2B可知ECM受体相互作用(ECM-receptor interaction)、补体和凝血级联反应(Complement and coagulation cascades)、癌症通路(Pathways in cancer)和AGE-RAGE信号通路(AGE-RAGE signaling pathway )等与环境污染物影响DN的病理生理变化关系密切.
中性粒细胞作为白细胞的重要组成部分,是机体炎症和感染的第一反应者,在人体天然和适应性免疫反应中发挥趋化、吞噬、脱颗粒等作用[19]. 当机体组织发生炎症反应时,发炎组织处中性粒细胞产生募集级联反应,在趋化因子的作用下黏附于血管壁,然后穿过血液—内皮细胞(endothelial cell,EC)屏障移行至发炎组织发挥细胞功能[20],中性粒细胞还可以通过提供基质重塑酶、形成中性粒细胞胞外陷阱(neutrophil extracellular traps,NETs)和释放外泌体等方式介导细胞外基质(extracellular matrix,ECM)重塑,反之ECM也可以通过调节中性粒细胞功能重塑炎症微环境[21]. 另外,活化的中性粒细胞在形成NETs的同时还会释放VEGFA、CXCL8、HGF和MMP9等血管生成因子,发挥原血管生成活性增加血管生成[22],并且NETs还参与补体级联反应和血小板激活,在形成血液高凝状态的同时促进肿瘤的发生及转移[23].
DN的发病机制十分复杂,炎症反应、氧化应激和血管血流动力学改变等因素都对DN发生发展影响巨大[24-25]. 研究表明,除IL-6、TNF-α等先天免疫系统效应分子水平在DN患者体内增高外[26],在肾脏活检中也发现了中性粒细胞的浸润,并且高血清中性粒细胞已经被证实与DN有关[27]. 虽然中心粒细胞微粒可以抑制IFN-γ和TNF-α的产生,但其增强了IL-2 / IL-12激活的自然杀伤细胞(natural killer cell,NK)释放TGF-β1[28],TGF-β1作为重要的致纤维化因子可诱发伴有肾纤维化和肾小球硬化的DN[29]. 除此之外,糖尿病及其并发症患者机体的高血糖状态还会诱导NETs的形成以及NETs产物的大量释放,进而导致NETosis[30]. 同时,高血糖机体环境形成的晚期糖基化终末产物(advanced glycation end products,AGEs)还可通过刺激循环多形核中性粒细胞产生活性氧(reactive oxygen species,ROS),与生长因子合成变化、炎症反应等一同造成血管内皮功能障碍,使DN患者产生蛋白尿[31],造成患者机体状况的进一步恶化.
环境污染物也可通过中性粒细胞途径影响机体健康. 暴露于合成内分泌干扰污染物比如双酚A等可直接影响人体免疫细胞的发育,影响免疫调节功能[32],2,2,4,4-四氯联苯可改变人粒细胞HL-60功能并增强其脱颗粒[33],甲基汞可以促发白细胞黏附并增加趋化因子表达[34]. 同时,环境污染物之间还存在协同效应,砷和乙醇的共暴露可诱导血管生成基因及VEGFA的表达[35],在调控血管生成的同时造成血流动力学的改变. 并且对于长期暴露个人而言,全氟辛酸的血清浓度水平与肾癌发生率呈正相关[36].
综上,环境污染物暴露可通过多种方式间接影响DN发生发展,并有明显的促癌倾向. 其中,以中性粒细胞为核心的一系列免疫炎症反应是机体暴露于环境污染物的主要生理活动,也是环境污染物影响DN并诱发癌症的生物学基础.
-
为了明确共同关联基因间的相互作用关系及挖掘影响共同基因的主要环境污染物,将1176个共同关联基因导入String数据库进行蛋白互作分析,隐藏断开节点的基因,共得到1154个节点,11913条边,代表了1154个共同关联基因通过11913条边相互联系,将其映射至Cytoscape软件,利用CytoHubba插件中的MCC算法筛选出Top200的基因作为关键基因进行后续分析,取Top20关键基因进行PPI网络可视化,如图3所示,颜色越深代表该基因在关键基因排序中越靠前,即在环境污染物影响DN发展进程中越重要.
-
利用Metascape在线分析平台对Top200基因进行Mcode模块化分析,显示有7个基因模块与环境污染物影响DN关系密切(图4),其中Mcode1最复杂,包含61个节点基因,远多于其他模块,将节点基因>5的模块及其所关联的环境污染物进行展示(表2),污染物按所关联节点基因的多少进行排序,污染物越靠前则代表其涉及的节点基因越多,即在环境污染物影响DN病理生理变化中越重要,是为主要环境污染物.
经过对Mcode1—4污染物进行联合分析,得出雌二醇、双酚A、镉、四氯二苯并二噁英、砷、尼古丁、颗粒物、多氯联苯等环境污染物可通过节点基因影响DN,内分泌干扰物雌二醇、双酚A是影响DN的主要环境污染物.
环境内分泌干扰物是指可以干扰人体激素正常生理过程及功能的环境污染物质,它们可以存在于空气、饮水和食物中并通过皮肤、呼吸、饮食等各种途径进入人体,起到类激素作用[37]. 环境内分泌干扰物种类繁多,包括雌二醇、双酚A、多氯联苯化合物、重金属等,因其大多数具有亲脂性,故非常容易在生物体内富集,影响正常机体激素稳态平衡和调节过程[38]. 雌二醇可通过甲基化等复杂表观遗传学机制影响基因表达调控,其整个过程与雌激素受体α关系密切[39]. 一项来自我国西北农村妇女的横断面研究发现,代谢综合征与女性机体血清雌二醇水平之间存在正相关,且与血清维生素D存在潜在协同作用[40]. 双酚A与雌二醇结构相似,广泛存在于塑料制品中,带来了极大的环境暴露威胁. Cimmino等[41]的研究表明,双酚A可通过影响胰岛素稳态和胰岛素信号传导的相关基因差异miRNA表达影响机体代谢,还可以通过DNA甲基化、组蛋白修饰等持久影响基因表达,这种持久的变化会导致神经免疫疾病、不孕不育症、迟发型复杂疾病(癌症和糖尿病)等一系列损伤,并可能产生跨代的遗传学毒性[42]. 与正常2型糖尿病患者相比,血清高水平双酚A的患者其发生DN的风险增加了7倍[43],而且双酚A还会引起足细胞减少、肾小球塌陷等类似糖尿病的早期肾脏改变[44],这无疑肯定了双酚A对DN的促进作用. 此外,金属镉、砷和四氯二苯并二恶英等环境污染物也可增加人糖尿病患病率,并且其暴露水平与患病严重程度正相关[45-47],同时,镉还可积聚在人近端肾小管处,通过影响线粒体电子传递链和NADPH氧化酶功能产生ROS造成核酸、蛋白质和脂质过氧化,在糖尿病高血糖的机体环境下加重肾脏损伤[48-49]. 再者,尼古丁和细颗粒物(particulate matter2.5, PM2.5)也被证实与慢性肾脏疾病进展密切相关[50-51],它们两者均可降低肾小球滤过率使尿白蛋白水平增加,并且尼古丁作为香烟烟雾的主要有害成分,可增加肾脏系膜细胞增殖和纤连蛋白产生促进肾脏纤维化,进而加重DN严重程度[52-54]. 综上,多种环境污染物均可对机体健康造成损伤,并在增加人糖尿病患病率的同时直接影响肾脏器官功能,协同机体高血糖状况加速DN恶化,而雌二醇和双酚A作为影响DN的主要环境污染物,其几乎贯穿内分泌及代谢性疾病始末,从基因乃至器官层面影响DN发生发展.
-
主要环境污染物与蛋白的分子对接结合情况如表3所示. 可知环境污染物雌二醇与双酚A与关键蛋白结合强度均<−4.5 kcal·mol−1,具有较强结合活性. 再者,分子对接还发现关键蛋白通过LEU、LYS、CYS、ARG、ASP、ILE和TYR等氨基酸残基形成氢键作用力与环境污染物相结合,且LEU和LYS出现频次最高,因此推测LEU和LYS是环境污染物雌二醇、双酚A与关键基因蛋白结合的主要残基. 选取结合最强的6个关键基因进行可视化(图5),位于结合图中间的青蓝色结构代表环境污染物,其他部位为与蛋白的结合位点.
-
将Top20关键基因导入在线分析平台GEPIA,分析其在KIRC组织中的表达水平. 经分析发现,Top20关键基因中有VEGFA、FN1、MMP9、PECAM1、CD44、ICAM1、CXCR4、TIMP1和PTPRC显著上调表达,EGF和CXCL12显著下调表达(图6),提示环境污染物暴露可能会增加DN本体罹患肾癌的风险.
-
对Top20关键基因进行Kaplan-Meier生存分析,生存率的比较采用Log-rank检验,结果如图7所示. 在Kaplan-Meier Plotter的677个KIRC病例中,结合基因表达谱数据动态分析结果,高表达的FN1、MMP9、PECAM1、CD44、ICAM1、CXCR4、TIMP1和低表达的CXCL12与KIRC患者的OS显著相关(P<0.05).
研究表明,与非糖尿病患者相比,糖尿病可显著增加糖尿病患者患肾癌的风险[55],并且造成的癌症死亡率超非糖尿病患者1倍以上[56]. 但目前关于DN与肾癌的关系尚不清楚,Luo等[57]的研究表明,DN大鼠钙调节的紊乱与其患肾癌的易感性有关. Dong[58]等通过生物信息学对DN和肾癌的共表达差异基因进行分析,发现DN和肾癌之间存在显著相关性,且FN1可能是DN相关肾癌的生物标志物之一. 这与本文的研究结果相符,并且既往研究也表明,FN1、MMP9、PECAM1、CD44、ICAM1、CXCR4、TIMP1和CXCL12这些关键基因表达水平都与KIRC预后相关,是诊断KIRC进展及预后的生物标志物[59-63],这也就证实了DN与肾癌之间确实存在相关性. 另外,对环境污染物暴露诱发肾癌的研究也表示,环境雌二醇暴露将增强癌症进展过程中CD44的表达[64],而双酚A则可以通过下调MMP9和上调TIMP1抑制细胞迁移和侵袭[65]. 镉的作用则恰巧相反,Shi[66]等的研究表明镉暴露可通过cAMP / PKA-COX2途径诱导肾癌细胞侵袭和迁移. 再者,长期暴露于无机砷则会诱发肾干细胞癌症表型,持续依赖性的表达MMP2和MMP9[67],同时尼古丁衍生物也可以剂量依赖性方式促进肾癌细胞生长和迁移[68]. 环境污染物与肾癌发生及癌细胞扩散关系密切,环境污染物影响DN的关键基因同样在肾癌发生中发挥作用,这也说明了环境污染物、DN和肾癌三者之间存在相互作用关系.
-
综上所述,本研究发现环境污染物暴露对DN发生发展影响巨大,并且还可能增加DN患者罹患肾癌的风险. 环境污染物在影响DN差异基因表达的同时,还可以通过以中心粒细胞为核心的脱颗粒、黏附、趋化和凝血等炎症反应、血管生成调节及癌症相关通路推动DN发展. 同时,还发现雌二醇和双酚A类环境内分泌干扰物是影响DN的主要环境污染物,它们主要通过LEU和LYS等氨基酸残基形成氢键作用力直接作用于非雌激素受体蛋白(关键基因). 最后,通过基因表达动态图谱和生存分析探讨了DN与肾癌之间的关系,而环境污染物的暴露无疑是增加了DN合并肾癌的机率. 本文为进一步研究环境污染物暴露影响DN提供了理论支持,可为临床DN精准防治及环境健康研究提供一定参考.
基于生物信息学分析研究环境污染物对糖尿病肾病的影响及其分子机制
Bioinformatics analysis of effects and molecular mechanism of environmental pollutants on diabetic nephropathy
-
摘要: 环境污染物是包括重金属、持久性有机污染物等在内一系列物质的总称,其可通过各种途径影响代谢类疾病及并发症的发生发展. 糖尿病肾病(DN)作为糖尿病最严重的微血管并发症之一,现已成为我国终末期肾病的主要病因. 本研究旨在探讨可作用于DN疾病变化进程的环境污染物及其调控机制,从而为糖尿病及DN患者精准预防和临床治疗提供一定理论依据. 通过Gene Expression Omnibus(GEO)和Comparative Toxicogenomics Database(CTD)数据库获取环境污染物影响DN的相关基因,其中经GEO数据库挖掘得到9个与DN相关联的数据集,标准化后筛选去重得到差异基因1326个,CTD数据库获得环境污染物影响DN的相关基因19557个,二者取交集得到共同基因1176个,利用DAVID数据库进行GO和KEGG富集分析,再将共同基因导入String数据库和Cytoscape软件获取关键基因,利用Metascape在线平台进行Mcode模块化分析,分析得出内分泌干扰物雌二醇、双酚A是影响DN的主要环境污染物,分子对接表明主要环境污染物与关键基因翻译后的蛋白结构结合活性良好,且主要通过LEU、LYS等氨基酸残基位点直接作用于关键基因蛋白. 最后,利用GEPIA在线平台验证Top20关键基因在肾透明细胞癌(KIRC)样本中的表达,并进行了Kaplan-Meier生存分析,其中8个显著表达基因与KIRC患者预后关系紧密,进一步探讨了环境污染物暴露介导的DN与肾癌关系. 综上,环境污染物可通过以中性粒细胞为核心的炎症反应诱发和促进DN,在影响DN病理生理进程的同时还对其合并肾癌存在诱发效应.Abstract: Environmental pollutants include heavy metals,persistent organic pollutants and so on,which can affect the occurrence and development of metabolic diseases and complications through various ways. Diabetic nephropathy (DN) is one of the most serious microvascular complications of diabetes,which has become the main cause of end-stage renal disease in China. This study aim to explore the environmental pollutants which play important role on the process of DN disease and its regulatory mechanisms its regulatory mechanisms,thus providing theoretical basis for the precise prevention and future clinical treatment of diabetes and DN patients. The related genes of environmental pollutants affecting DN were obtained through Gene Expression Omnibus(GEO)and Comparative Toxicogenomics Database(CTD)databases. Then,9 datasets associated with DN were obtained from the GEO database. After standardization,1326 differential genes were obtained by screening and deduplication. 19557 genes related to environmental pollutants affecting DN were obtained from the CTD database,and 1176 common genes were obtained between GEO and CTD database. Next,GO and KEGG enrichment analyses were conducted through DAVID database,and then the common genes were imported into String database and Cytoscope software to obtain key genes. Mcode modular analysis were performed using the Metascape online platform. The analysis showed that the endocrine disruptors estradiol and bisphenol A were the main environmental pollutants affecting DN. Molecular docking showed that the main environmental pollutants had strong binding activity with proteins structure of key genes,and they mainly acted directly through amino acid residues such as LEU and LYS. Finally,the expression of these Top20 key genes in kidney renal clear cell carcinoma (KIRC) sample was further verified using GEPIA online platform and Kaplan-Meier survival curves were constructed.The results showed that 7 significantly expressed genes were closely related to the prognosis of KIRC patients,and further explored the relationship between DN and renal cancer mediated by environmental pollutants exposure. In conclusion, environmental pollutants can induce and promote DN through the inflammatory response and neutrophils play an extremely important role in these process. Further,it also has an inducing effect on its concomitant kidney cancer while affecting the pathophysiological process of DN.
-
Key words:
- environmental pollutants /
- diabetic nephropathy /
- bioinformatics /
- estradiol /
- bisphenol A.
-

-
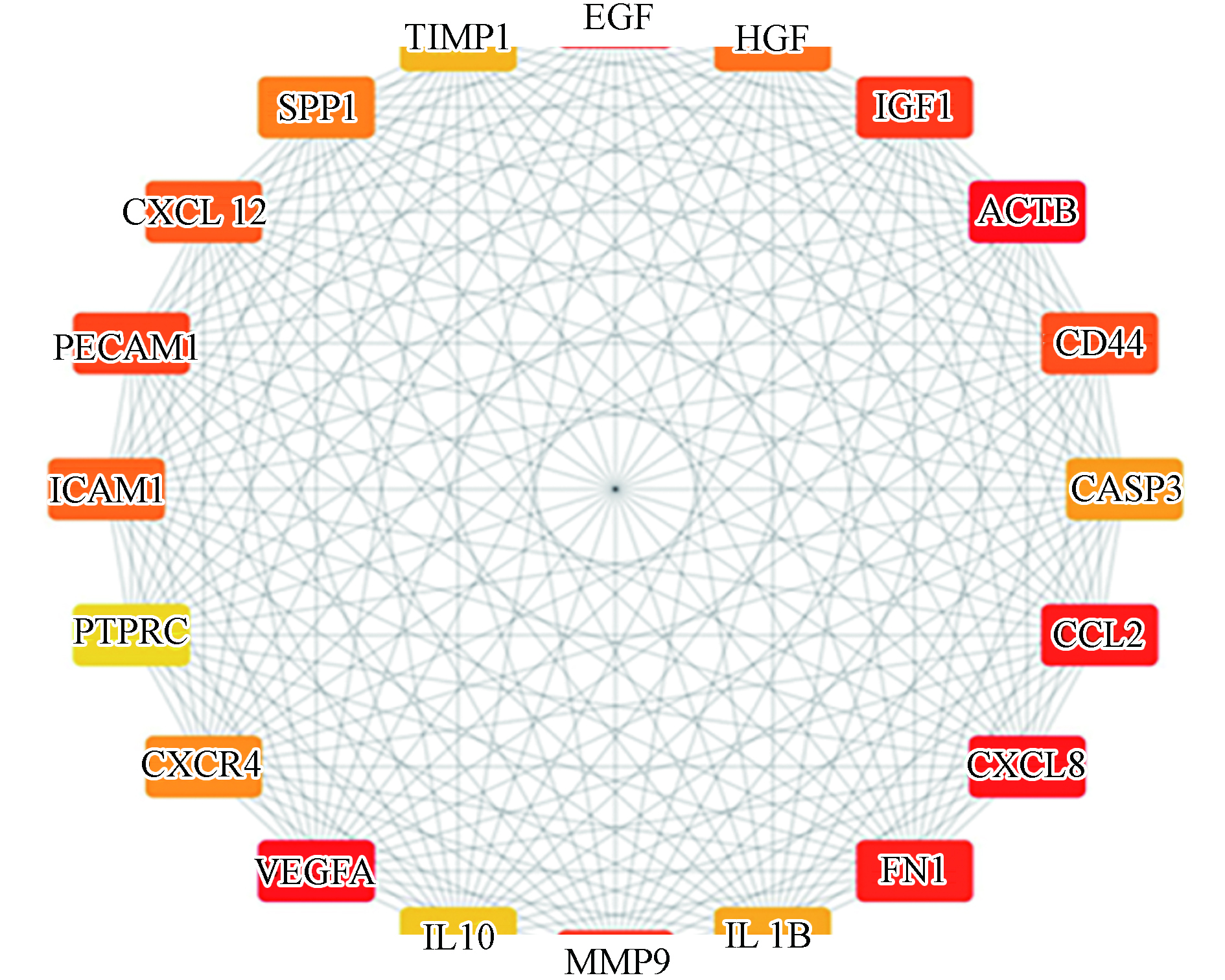
图 3 Top20关键基因蛋白相互作用网络图
Figure 3. The protein -protein interaction network of top20 key genes

图 4 基因模块图及主要环境污染物
Figure 4. The module diagram of genes and main environmental pollutants
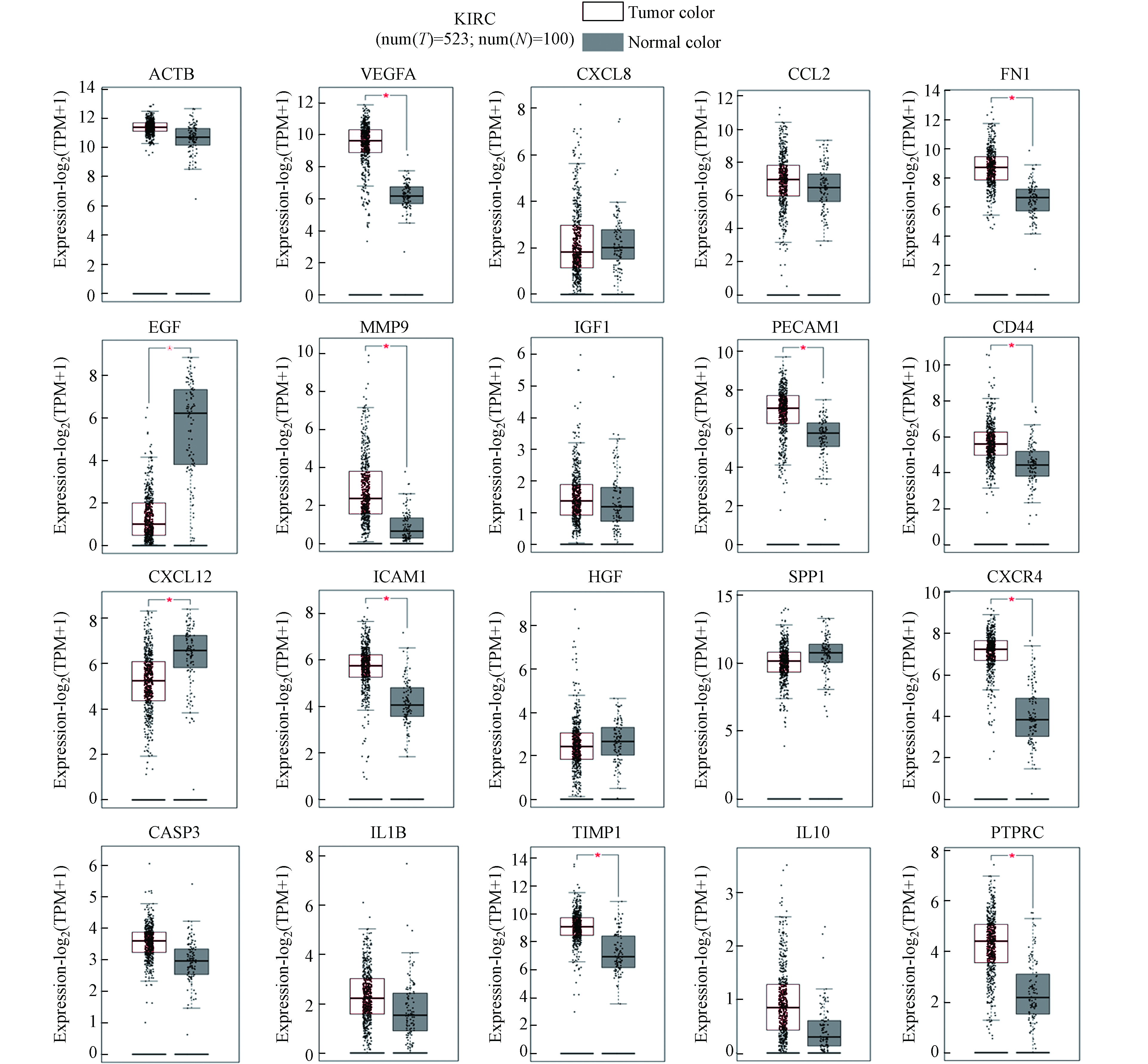
图 6 Top20关键基因在KIRC组织中的相对表达水平比较(*P<0.05)
Figure 6. Comparison of relative expression levels of Top20 key genes in Kidney renal clear cell carcinoma tissue(*P<0.05)

图 7 Top20关键基因表达与KIRC患者OS的关系
Figure 7. Association between Top20 key genes expression and OS of Kidney renal clear cell carcinoma patients
表 1 DN相关基因芯片数据集信息
Table 1. DN related gene chip datasets information
GSE登记号
Accession国家
Country取材组织
Sampling tissue年龄范围
Age性别及例数
Gebder and numberDN样本
DN sample非DN样本
Non-DN sampleGSE1009 荷兰 肾小球 29—70 4男 2 2 GSE142153 美国 外周血 26—71 19男/14女 23 10 GSE4117 日本 外周血 56—71 5男/2女 3 4 GSE47185 美国 GSE47183 GPL14663 肾组织 — — 7 61 GSE47184 GPL14663 肾组织 — — 11 53 GSE30122 美国 GSE30528 肾小球 39—78 12男/10女 9 13 GSE30529 肾小管 40—80 8男/14女 10 12 GSE46900 美国 GSE46899 足细胞 33—58 6男 2 4 GSE46897 足细胞 33—58 6男 2 4  下载: 导出CSV
下载: 导出CSV
表 2 影响Mcode模块的环境污染物
Table 2. Environmental pollutants affecting the Mcode module
模块
Mcode关键基因
Key genes污染物
PollutantsMcode1 CCR2、IFITM2、IFITM3、IFITM1、ITGA8、CXCR4、VWF、VCAM1、CXCL12、CXCL6、CCL20、CCL19、CCL5、PSMB8、MX1、CXCL9、KNG1、ITGB8、ITGB5、ITGB3、ITGB1、ITGAV、ITGA4、ITGA3、ISG20、IRF1、CXCL8、IFIT1、IRF8、ICAM1、HLA-F、HLA-E、HLA-DRB5、HLA-DRB1、HLA-DRA、HLA-DQB1、HLA-DPB1、HLA-DPA1、HLA-C、HLA-B、HLA-A、CXCL3、CXCL2、CXCL1、GBP2、IFI6、FN1、CX3CR1、COL6A3、COL5A2、COL4A6、COL4A5、COL4A3、COL4A2、COL4A1、COL3A1、COL1A2、CD44、C3、BST2、ANXA1 雌二醇、镉、双酚A、 四氯二苯并二噁英、 砷、颗粒物、铜、钴、镍、2,4,5,2',4',5'-六氯联苯、2,5,2',5'-四氯联苯、苯并芘、过氧化氢、石棉、车辆排放物、锌 Mcode2 VEGFA、CCL2、KDR、JAK1、IL10、IGF1、FYN、FLT1、EGF、VCAN、CSF2RB、CAV1 雌二醇、双酚A、镉、四氯二苯并二噁英、 砷、3,4,5,3',4'-五氯联苯、2,4,5,2',4',5'-六氯联苯、颗粒物、对苯二酚、尼古丁、苯并芘、锌 、石棉、钠 Mcode3 THBS2、THBS1、TGFBI、PTPRC、MMP9、LYN、LCP2、LCK、ITGB2、ITGAM、CD3D 雌二醇、双酚A、四氯二苯并二噁英、镉、钴、铁、尼古丁、砷、邻苯二甲酸二乙基己酯、3,4,5,3',4'-五氯联苯 Mcode4 SYK、MYCIKBKB、FAS、ANXA5、ANXA2、ACTB 雌二醇、氯化钴、尼古丁、钴、铁、过氧化氢、棕榈油
下载: 导出CSV
表 3 分子对接结合情况
Table 3. Molecular docking binding
污染物
Pollutants关键靶点
Key targets结合能/(kcal·mol−1)
Binding energy污染物
Pollutants关键靶点
Key targets结合能/(kcal·mol−1)
Binding energyEstradiol ACTB −6.19 Bisphenol A ACTB −5.22 VEGFA −6.65 VEGFA −6.08 CXCL8 −8.13 CXCL8 −7.41 CCL2 −6.72 CCL2 −5.47 FN1 −6.5 FN1 −4.59 EGF −6.99 EGF −6.69 MMP9 −7.62 MMP9 −7.5 IGF1 −6.54 IGF1 −5.17 PECAM1 −6.12 PECAM1 −5.21 CD44 −10.08 CD44 −6.86 CXCL12 −5.18 CXCL12 −4.59 ICAM1 −6.55 ICAM1 −5.9 HGF −7.82 HGF −5.36 SPP1 −7.14 SPP1 −6.54 CXCR4 −6.31 CXCR4 −5.06 CASP3 −6.96 CASP3 −6.17 IL1B −6.93 IL1B −5.21 TIMP1 −6.69 TIMP1 −4.52 IL10 −7.18 IL10 −5.45 PTPRC −5.69 PTPRC −5.02
下载: 导出CSV
-
[1] WANG F, YANG C, LONG J Y, et al. Executive summary for the 2015 annual data report of the China kidney disease network (CK-NET) [J]. Kidney International, 2019, 95(3): 501-505. doi: 10.1016/j.kint.2018.11.011 [2] DOMINGUETI C P, DUSSE L M S, CARVALHO M D G, et al. Diabetes mellitus: The linkage between oxidative stress, inflammation, hypercoagulability and vascular complications [J]. Journal of Diabetes and Its Complications, 2016, 30(4): 738-745. doi: 10.1016/j.jdiacomp.2015.12.018 [3] SAGOO M K, GNUDI L. Diabetic nephropathy: An overview [J]. Methods in Molecular Biology, 2020, 2067: 3-7. [4] LIU D W, LI Z Y, LIU Z S. Treatment of diabetic kidney disease: Research development, current hotspots and future directions [J]. Zhonghua Yi Xue Za Zhi, 2021, 101(10): 683-686. [5] BONNER R, ALBAJRAMI O, HUDSPETH J, et al. Diabetic kidney disease [J]. Primary Care:Clinics in Office Practice, 2020, 47(4): 645-659. doi: 10.1016/j.pop.2020.08.004 [6] GÜLÜMSEK E, KEŞKEK Ş Ö. Direct medical cost of nephropathy in patients with type 2 diabetes [J]. International Urology and Nephrology, 2022, 54(6): 1383-1389. doi: 10.1007/s11255-021-03012-4 [7] HOU Y, GAO Y, ZHANG Y, et al. Interaction between ELMO1 gene polymorphisms and environment factors on susceptibility to diabetic nephropathy in Chinese Han population [J]. Diabetology & Metabolic Syndrome, 2019, 11: 97. [8] LIU L, WU Q C, MIAO X Y, et al. Study on toxicity effects of environmental pollutants based on metabolomics: A review [J]. Chemosphere, 2022, 286: 131815. doi: 10.1016/j.chemosphere.2021.131815 [9] HUANG W H, LIN J L, LIN-TAN D T, et al. Environmental lead exposure accelerates progressive diabetic nephropathy in type II diabetic patients [J]. BioMed Research International, 2013, 2013: 742545. [10] GRICE B A, NELSON R G, WILLIAMS D E, et al. Associations between persistent organic pollutants, type 2 diabetes, diabetic nephropathy and mortality [J]. Occupational and Environmental Medicine, 2017, 74(7): 521-527. doi: 10.1136/oemed-2016-103948 [11] EVERETT C J, THOMPSON O M. Dioxins, furans and dioxin-like PCBs in human blood: Causes or consequences of diabetic nephropathy? [J]. Environmental Research, 2014, 132: 126-131. doi: 10.1016/j.envres.2014.03.043 [12] GONG P, WANG P P, PI S H, et al. Proanthocyanidins protect against cadmium-induced diabetic nephropathy through p38 MAPK and Keap1/Nrf2 signaling pathways [J]. Frontiers in Pharmacology, 2022, 12: 801048. doi: 10.3389/fphar.2021.801048 [13] LAWRENCE S M, CORRIDEN R, NIZET V. The ontogeny of a neutrophil: Mechanisms of granulopoiesis and homeostasis [J]. Microbiology and Molecular Biology Reviews:MMBR, 2018, 82(1): e00057-e00017. [14] TSAI H J, HUNG C H, WANG C W, et al. Associations among heavy metals and proteinuria and chronic kidney disease [J]. Diagnostics (Basel, Switzerland), 2021, 11(2): 282. [15] RAMHARACK P, SOLIMAN M E S. Bioinformatics-based tools in drug discovery: The cartography from single gene to integrative biological networks [J]. Drug Discovery Today, 2018, 23(9): 1658-1665. doi: 10.1016/j.drudis.2018.05.041 [16] VAN J A D, SCHOLEY J W, KONVALINKA A. Insights into diabetic kidney disease using urinary proteomics and bioinformatics [J]. Journal of the American Society of Nephrology:JASN, 2017, 28(4): 1050-1061. doi: 10.1681/ASN.2016091018 [17] TZIASTOUDI M, CHOLEVAS C, THEOHARIDES T C, et al. Meta-analysis and bioinformatics detection of susceptibility genes in diabetic nephropathy [J]. International Journal of Molecular Sciences, 2021, 23(1): 20. doi: 10.3390/ijms23010020 [18] VERGA J U, HUFF M, OWENS D, et al. Integrated genomic and bioinformatics approaches to identify molecular links between endocrine disruptors and adverse outcomes [J]. International Journal of Environmental Research and Public Health, 2022, 19(1): 574. doi: 10.3390/ijerph19010574 [19] RAVICHANDRAN J, KARTHIKEYAN B S, APARNA S R, et al. Network biology approach to human tissue-specific chemical exposome [J]. The Journal of Steroid Biochemistry and Molecular Biology, 2021, 214: 105998. doi: 10.1016/j.jsbmb.2021.105998 [20] FILIPPI M D. Neutrophil transendothelial migration: Updates and new perspectives [J]. Blood, 2019, 133(20): 2149-2158. doi: 10.1182/blood-2018-12-844605 [21] ZHU Y Y, HUANG Y M, JI Q, et al. Interplay between extracellular matrix and neutrophils in diseases [J]. Journal of Immunology Research, 2021, 2021: 8243378. [22] POTO R, CRISTINZIANO L, MODESTINO L, et al. Neutrophil extracellular traps, angiogenesis and cancer [J]. Biomedicines, 2022, 10(2): 431. doi: 10.3390/biomedicines10020431 [23] MASUCCI M T, MINOPOLI M, del VECCHIO S, et al. The emerging role of neutrophil extracellular traps (NETs) in tumor progression and metastasis [J]. Frontiers in Immunology, 2020, 11: 1749. doi: 10.3389/fimmu.2020.01749 [24] FEIGERLOVÁ E, BATTAGLIA-HSU S F. IL-6 signaling in diabetic nephropathy: From pathophysiology to therapeutic perspectives [J]. Cytokine & Growth Factor Reviews, 2017, 37: 57-65. [25] GEWIN L S. TGF-β and diabetic nephropathy: Lessons learned over the past 20 years [J]. The American Journal of the Medical Sciences, 2020, 359(2): 70-72. doi: 10.1016/j.amjms.2019.11.010 [26] CHEN Y L, QIAO Y C, XU Y, et al. Serum TNF-α concentrations in type 2 diabetes mellitus patients and diabetic nephropathy patients: A systematic review and meta-analysis [J]. Immunology Letters, 2017, 186: 52-58. doi: 10.1016/j.imlet.2017.04.003 [27] WINTER L, WONG L A, JERUMS G, et al. Use of readily accessible inflammatory markers to predict diabetic kidney disease [J]. Frontiers in Endocrinology, 2018, 9: 225. doi: 10.3389/fendo.2018.00225 [28] PLIYEV B K, KALINTSEVA M V, ABDULAEVA S V, et al. Neutrophil microparticles modulate cytokine production by natural killer cells [J]. Cytokine, 2014, 65(2): 126-129. doi: 10.1016/j.cyto.2013.11.010 [29] HORIKOSHI S, FUKUDA N, TSUNEMI A, et al. Contribution of TGF-β1 and effects of gene silencer pyrrole-imidazole polyamides targeting TGF-β1 in diabetic nephropathy [J]. Molecules, 2020, 25(4): 950. doi: 10.3390/molecules25040950 [30] NJEIM R, AZAR W S, FARES A H, et al. NETosis contributes to the pathogenesis of diabetes and its complications [J]. Journal of Molecular Endocrinology, 2020, 65(4): R65-R76. doi: 10.1530/JME-20-0128 [31] PATHOMTHONGTAWEECHAI N, CHUTIPONGTANATE S. AGE/RAGE signaling-mediated endoplasmic reticulum stress and future prospects in non-coding RNA therapeutics for diabetic nephropathy [J]. Biomedicine & Pharmacotherapy, 2020, 131: 110655. [32] NOWAK K, JABŁOŃSKA E, RATAJCZAK-WRONA W. Immunomodulatory effects of synthetic endocrine disrupting chemicals on the development and functions of human immune cells [J]. Environment International, 2019, 125: 350-364. doi: 10.1016/j.envint.2019.01.078 [33] BEZDECNY S A, ROTH R A, GANEY P E. Effects of 2, 2', 4, 4'-tetrachlorobiphenyl on granulocytic HL-60 cell function and expression of cyclooxygenase-2 [J]. Toxicological Sciences, 2005, 84(2): 328-334. doi: 10.1093/toxsci/kfi093 [34] FOWLER J, TSUI M T K, CHAVEZ J, et al. Methyl mercury triggers endothelial leukocyte adhesion and increases expression of cell adhesion molecules and chemokines [J]. Experimental Biology and Medicine , 2021, 246(23): 2522-2532. doi: 10.1177/15353702211033812 [35] KLEI L R, BARCHOWSKY A. Positive signaling interactions between arsenic and ethanol for angiogenic gene induction in human microvascular endothelial cells [J]. Toxicological Sciences, 2008, 102(2): 319-327. doi: 10.1093/toxsci/kfn003 [36] SHEARER J J, CALLAHAN C L, CALAFAT A M, et al. Serum concentrations of per- and polyfluoroalkyl substances and risk of renal cell carcinoma [J]. JNCI:Journal of the National Cancer Institute, 2020, 113(5): 580-587. [37] 李秀婷, 王军, 赵亮亮, 等. 环境内分泌干扰物与糖尿病发病关联的研究进展 [J]. 环境与健康杂志, 2018, 35(5): 465-469. doi: 10.16241/j.cnki.1001-5914.2018.05.024 LI X T, WANG J, ZHAO L L, et al. Environmental endocrine disruptors and diabetes: A review of recent studies [J]. Journal of Environment and Health, 2018, 35(5): 465-469(in Chinese). doi: 10.16241/j.cnki.1001-5914.2018.05.024
[38] 王航, 张李一, 张蕴晖. 主要环境内分泌干扰物疾病负担的研究进展 [J]. 环境与职业医学, 2021, 38(9): 1033-1043. doi: 10.13213/j.cnki.jeom.2021.20565 WANG H, ZHANG L Y, ZHANG Y H. Research progress on disease burdens of major environmental endocrine disruptors [J]. Journal of Environmental and Occupational Medicine, 2021, 38(9): 1033-1043(in Chinese). doi: 10.13213/j.cnki.jeom.2021.20565
[39] KOVÁCS T, SZABÓ-MELEG E, ÁBRAHÁM I M. Estradiol-induced epigenetically mediated mechanisms and regulation of gene expression [J]. International Journal of Molecular Sciences, 2020, 21(9): 3177. doi: 10.3390/ijms21093177 [40] YANG C, LIU X W, LI J, et al. Association of serum vitamin D and estradiol levels with metabolic syndrome in rural women of northwest China: A cross-sectional study [J]. Metabolic Syndrome and Related Disorders, 2022, 20(3): 182-189. doi: 10.1089/met.2021.0120 [41] CIMMINO I, FIORY F, PERRUOLO G, et al. Potential mechanisms of bisphenol A (BPA) contributing to human disease [J]. International Journal of Molecular Sciences, 2020, 21(16): 5761. doi: 10.3390/ijms21165761 [42] SINGH S, LI S S L. Epigenetic effects of environmental chemicals bisphenol A and phthalates [J]. International Journal of Molecular Sciences, 2012, 13(8): 10143-10153. doi: 10.3390/ijms130810143 [43] HU J B, YANG S M, WANG Y, et al. Serum bisphenol A and progression of type 2 diabetic nephropathy: A 6-year prospective study [J]. Acta Diabetologica, 2015, 52(6): 1135-1141. doi: 10.1007/s00592-015-0801-5 [44] MORENO-GÓMEZ-TOLEDANO R, ARENAS M I, MUÑOZ-MORENO C, et al. Comparison of the renal effects of bisphenol A in mice with and without experimental diabetes. Role of sexual dimorphism [J]. Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease, 2022, 1868(1): 166296. doi: 10.1016/j.bbadis.2021.166296 [45] EDWARDS J R, PROZIALECK W C. Cadmium, diabetes and chronic kidney disease [J]. Toxicology and Applied Pharmacology, 2009, 238(3): 289-293. doi: 10.1016/j.taap.2009.03.007 [46] GOODMAN M, NARAYAN K M V, FLANDERS D, et al. Dose-response relationship between serum 2, 3, 7, 8-tetrachlorodibenzo-p-dioxin and diabetes mellitus: A Meta-analysis [J]. American Journal of Epidemiology, 2015, 181(6): 374-384. doi: 10.1093/aje/kwu307 [47] FARKHONDEH T, SAMARGHANDIAN S, AZIMI-NEZHAD M. The role of arsenic in obesity and diabetes [J]. Journal of Cellular Physiology, 2019, 234(8): 12516-12529. doi: 10.1002/jcp.28112 [48] YAN L J, ALLEN D C. Cadmium-induced kidney injury: Oxidative damage as a unifying mechanism [J]. Biomolecules, 2021, 11(11): 1575. doi: 10.3390/biom11111575 [49] LI M Y, LIU X X, ZHANG Z L. Hyperglycemia exacerbates cadmium-induced glomerular nephrosis[J]. Toxicology and Industrial Health, 2021, 37(4): 074823372110378. . [50] JAIMES E A, ZHOU M S, SIDDIQUI M, et al. Nicotine, smoking, podocytes, and diabetic nephropathy [J]. American Journal of Physiology. Renal Physiology, 2021, 320(3): F442-F453. doi: 10.1152/ajprenal.00194.2020 [51] RASKING L, VANBRABANT K, BOVÉ H, et al. Adverse Effects of fine particulate matter on human kidney functioning: A systematic review [J]. Environmental Health:a Global Access Science Source, 2022, 21(1): 24. [52] LAN X Q, WEN H X, ASLAM R, et al. Nicotine enhances mesangial cell proliferation and fibronectin production in high glucose milieu via activation of Wnt/β-catenin pathway [J]. Bioscience Reports, 2018, 38(3): BSR20180100. doi: 10.1042/BSR20180100 [53] HUA P, FENG W G, JI S N, et al. Nicotine worsens the severity of nephropathy in diabetic mice: Implications for the progression of kidney disease in smokers [J]. American Journal of Physiology. Renal Physiology, 2010, 299(4): F732-F739. doi: 10.1152/ajprenal.00293.2010 [54] CHIN W S, CHANG Y K, HUANG L F, et al. Effects of long-term exposure to CO and PM2.5 on microalbuminuria in type 2 diabetes [J]. International Journal of Hygiene and Environmental Health, 2018, 221(4): 602-608. doi: 10.1016/j.ijheh.2018.04.009 [55] BAO C P, YANG X L, XU W L, et al. Diabetes mellitus and incidence and mortality of kidney cancer: A meta-analysis [J]. Journal of Diabetes and Its Complications, 2013, 27(4): 357-364. doi: 10.1016/j.jdiacomp.2013.01.004 [56] TSENG C H. Type 2 diabetes mellitus and kidney cancer risk: A retrospective cohort analysis of the national health insurance [J]. PLoS One, 2015, 10(11): e0142480. doi: 10.1371/journal.pone.0142480 [57] LUO Y M, LU Z Y, WAAGA-GASSER A M, et al. Modulation of calcium homeostasis may be associated with susceptibility to renal cell carcinoma in diabetic nephropathy rats [J]. Cancer Management and Research, 2020, 12: 9679-9689. doi: 10.2147/CMAR.S268402 [58] DONG Y Z, ZHAI W, XU Y F. Bioinformatic gene analysis for potential biomarkers and therapeutic targets of diabetic nephropathy associated renal cell carcinoma [J]. Translational Andrology and Urology, 2020, 9(6): 2555-2571. doi: 10.21037/tau-19-911 [59] YANG J F, SHI S N, XU W H, et al. Screening, identification and validation of CCND1 and PECAM1/CD31 for predicting prognosis in renal cell carcinoma patients [J]. Aging, 2019, 11(24): 12057-12079. doi: 10.18632/aging.102540 [60] KROEZE S G C, BIJENHOF A M, BOSCH J L H R, et al. Diagnostic and prognostic tissuemarkers in clear cell and papillary renal cell carcinoma [J]. Cancer Biomarkers:Section A of Disease Markers, 2010, 7(6): 261-268. [61] BERGLUND A, AMANKWAH E K, KIM Y C, et al. Influence of gene expression on survival of clear cell renal cell carcinoma [J]. Cancer Medicine, 2020, 9(22): 8662-8675. doi: 10.1002/cam4.3475 [62] WU F, WU S, GOU X. Identification of biomarkers and potential molecular mechanisms of clear cell renal cell carcinoma [J]. Neoplasma, 2018, 65(2): 242-252. doi: 10.4149/neo_2018_170511N342 [63] CHEN L, XIANG Z J, CHEN X R, et al. A seven-gene signature model predicts overall survival in kidney renal clear cell carcinoma [J]. Hereditas, 2020, 157(1): 38. doi: 10.1186/s41065-020-00152-y [64] YOUNG M J, CHEN Y C, WANG S A, et al. Estradiol-mediated inhibition of Sp1 decreases miR-3194-5p expression to enhance CD44 expression during lung cancer progression [J]. Journal of Biomedical Science, 2022, 29(1): 3. doi: 10.1186/s12929-022-00787-1 [65] WEI P, RU D Q, LI X Q, et al. Exposure to environmental bisphenol A inhibits HTR-8/SVneo cell migration and invasion [J]. Journal of Biomedical Research, 2020, 34(5): 369-378. doi: 10.7555/JBR.34.20200013 [66] SHI H F, SUN X, KONG A Q, et al. Cadmium induces epithelial-mesenchymal transition and migration of renal cancer cells by increasing PGE2 through a cAMP/PKA-COX2 dependent mechanism [J]. Ecotoxicology and Environmental Safety, 2021, 207: 111480. doi: 10.1016/j.ecoenv.2020.111480 [67] TOKAR E J, PERSON R J, SUN Y, et al. Chronic exposure of renal stem cells to inorganic arsenic induces a cancer phenotype [J]. Chemical Research in Toxicology, 2013, 26(1): 96-105. doi: 10.1021/tx3004054 [68] HUANG Y F, WANG Q Z, TANG Y, et al. Identification and validation of a cigarette smoke-related five-gene signature as a prognostic biomarker in kidney renal clear cell carcinoma [J]. Scientific Reports, 2022, 12: 2189. doi: 10.1038/s41598-022-06352-y -
 点击查看大图
点击查看大图
计量
- 文章访问数: 4131
- HTML全文浏览数: 4131
- PDF下载数: 174
- 施引文献: 0
