

-
环境化学品是指存在于空气、水、土壤、食物、灰尘或其他环境介质(如个人消费品等)中的人工合成或天然生成的化学物质(元素). 目前,在尿液、血液、血清、母乳和胎粪等人类样品中已检测到400余种环境化学品及其代谢产物(Centers for Disease Control and Prevention, CDC,
https://www.cdc.gov/biomonitoring /environmental_chemicals.html ). 环境化学品中多环芳烃(PAHs)等持久性有机污染物(POPs)、重金属、以及双酚类和抗生素等新污染物大多属于内分泌干扰物(EDCs),具有破坏内分泌功能的作用,可改变机体激素代谢,使机体内分泌紊乱. 环境化学品与人类生活息息相关,个人护理品、杀虫剂、消毒剂等的使用使其广泛暴露,引发机体微生态平衡失调,其短期急性或长期慢性暴露都可能引起许多不良的健康结局,包括肥胖症[1-2]、心血管疾病[3]、Ⅱ型糖尿病[4]、生殖发育毒性[5-6]、肝脏毒性[7-8]和肾脏毒性[9]等. 然而,由此诱导的不良健康结局也因不同的人群队列、年龄、性别等因素而有所差异(图1).越来越多的研究表明环境化学品暴露会改变宿主机体肠道微生物群组成,且大多研究表明肠道菌群组成变化与化学品暴露导致的不良健康影响相关(图1)[10-11]. 肠道微生物组是一个复杂的、高度动态的生态系统,不同个体间差异较大,其组成和功能很容易受到各种内源性和外源性因素的影响[12]. 不同基因型、同一个体的不同年龄段、不同地理位置和生活方式的、甚至单卵双胞胎的人或动物具有不同的肠道菌群[13-16]. 然而,异源物质与微生物组的关联通常是复杂的,具有双向相互作用. 一方面,环境化学品暴露可能会诱发宿主肠道微生物群组成和结构的改变,而微生物组组成的改变也会以产生代谢物等多种方式反馈给宿主,从而影响宿主机体代谢的改变,对宿主健康产生影响. 另一方面,肠道菌群也可以调节异源化学品暴露对宿主的毒性作用(图2). Younan等[17]认为,肠道菌群能够将高毒性的铬(Ⅵ)转化为更低毒性的铬(Ⅲ),另一项在黄鳍鲷中的研究发现,肠道中发生无机汞甲基化的现象,且特定的肠道菌群在这一过程中发挥重要作用,使无机汞转变成毒性更大的甲基汞[18]. 同时,在斑马鱼中通过添加鼠李糖乳杆菌可通过调节菌群多样性及结构来缓解全氟丁烷磺酸盐(PFBS)暴露引起的机体氧化应激和脂质代谢紊乱,从而减轻PFBS暴露的毒性作用,维持宿主健康[19].
本文总结了传统POPs、重金属和新污染物等三大类环境化学品暴露对宿主机体肠道菌群微生态的影响. 生物体长期暴露于各种环境化学品,了解肠道菌群如何响应环境化学品暴露及其肠道菌群变化对宿主健康的影响至关重要. 因此,本综述除了总结这些化学品暴露对宿主微生物组的影响外,还总结了与微生物组变化相关的基因表达、功能和代谢改变及其对宿主健康的影响.
-
POPs是一类具有持久性、生物累积效应、长距离迁移性和高毒性的有机化学物质. 因其难降解特性和亲脂性等特点,POPs易在生物体内富集并产生一定的健康风险[20]. 目前,越来越多的证据表明,POPs会破坏动物体内肠道微生物组的稳态[21]. 肠道微生物群是宿主的重要组成部分,负责许多重要的生理过程,包括能量和营养循环、肠道屏障维持及免疫系统的调节等[22] (表1). 因此,对POPs暴露后宿主肠道微生物组变化的分析可在一定程度上探究其毒性的作用方式和机制.
-
PAHs是碳质物质在不完全燃烧或热解过程中形成的,是环境中普遍存在的污染物[23]. 原油中含有大量PAHs类物质,例如萘、苯并[a]芘等,目前已有文章报道了海洋中鱼类通过吞食或皮肤、鳃吸收过量原油可对其自身造成多重致死效应,包括DNA损伤、心脏毒性、免疫抑制、生殖能力降低和肝脏肿瘤等[24, 25]. 除此之外,研究人员还注意到原油污染可能对海洋生物体内微生物造成一定的影响,如2016年加拿大赫斯基能源公司(Husky Energy)的重质原油污染事件中,漏油区域上下游中金眼鱼(Hiodon alosoides)、白眼鱼(Sander vitreus)、北梭子鱼(Esox lucius)和短头红马鱼(Moxostoma macrolepidotum)肌肉中PAHs含量与个体肠道微生物中的细菌丰度和组成呈现相关关系[26];而在2010年墨西哥湾“深水地平线”石油泄漏事件中,比目鱼(Paralichthys lethostigma)鳃和肠道中碳氢化合物代谢细菌Alcanivorax sp.丰度显著增加[27]. 轮胎颗粒(TPs)会向环境中释放PAHs和重金属等污染物,其可能通过影响土壤动物及其肠道微生物群以损害陆地生态系统的稳定性. 随着TPs浓度的增加,土壤中蠕虫E. crypticus存活率和繁殖率均有着不同程度的下降,同时TPs暴露使蠕虫肠道内与致病相关的微生物大量富集[28]. 此外 随着PAHs与空气污染间相关关系的逐渐证实,研究人员相继发现大气中PAHs污染可能改变与健康和环境相关的共生微生物群落,如:放线菌(Actinobacteria)丰度与大气中高分子量PAHs呈正相关,芽孢杆菌属和低分子量多环芳烃呈正相关、居住在森林附近的人体肠道微生物中芳香烃受体 (AhR) 激活受体更为活跃等[29-31].
而在众多PAHs中,苯并[a]芘(BaP)作为其中一种代表性物质,其对肠道微生物的影响及进一步造成的环境健康风险得到了深入的研究[21]. 在早期的一项研究中发现尖齿胡鲶(Clarias gariepinus)肠道中的Pseudomonas aeruginosa数量与其肌肉中注射的的BaP浓度相关,表明BaP会对肠道中特定微生物的丰度造成一定的影响[32]. 成年黑头鲦鱼和日本海参暴露在低浓度的BaP环境中会导致肠道中与烃类降解相关的微生物富集[33-34],也会造成年西方蚊鱼和斑马鱼的肠道微生物群失调并进一步产生炎症反应[35];除此之外,DeBofsky等[36]研究表明,在高浓度条件下(1000 μg·mg−1食物),BaP能显著降低微生物群落多样性、改变群落组成甚至在一定程度上降低微生物群落间的网络复杂性. 对于小鼠而言,当其口服 50 mg·kg−1(体重)BaP后,其肠道和粪便中有益A. muciniphila丰度增加,该菌可通过降解肠道的粘液和刺激肠上皮分泌丙酸的方式增强肠道屏障功能,但随着暴露时间的延长肠道内乳酸菌(Lactobacillus)、Mucispirillum、A. muciniphila等有益菌群丰度迅速下降,与此同时肠道中与炎症相关的Allobaculum和脱硫弧菌Desulfovibrionaceae数量有所上升,由此导致宿主回肠和结肠黏膜中度炎症的发生[37].
-
PCBs是一类典型的POPs和内分泌干扰物(EDCs),虽早已在20世纪被禁用,但至今仍能在一般人群的血清和母乳中检出,因此PCBs污染所引发的环境健康威胁一直是目前的研究重点. PCBs对生物体的毒性体现在诸多方面,如致癌性、生殖毒性、心血管炎症、甲状腺毒性以及肥胖、II型糖尿病和脂肪性肝病等[38].
PCBs是一种亲脂化合物,易于在脂肪组织中积累,相关研究证实即使暴露于低剂量也会导致肥胖,因此证明肥胖和PCBs暴露之间存在密切联系[39]. 与之相似,肠道微生物组组成的变化与多种代谢疾病相关,尤其是肥胖和糖尿病[22]. 早在2013年,Choi等[40]通过比较运动与不运动小鼠对PCBs暴露后菌群的影响研究发现,PCBs暴露降低小鼠肠道菌群丰度,而运动小鼠微生物丰富度和多样性有所提高,运动改变了PCBs对肠道菌群的介导作用,因此该实验表明小鼠能通过运动以减少PCBs暴露引起的微生物组的变化程度. 此后,大量文献相继报道了PCB77、PCB126等物质暴露所引起的肠道微生物特定种群的变化可在一定程度上削弱机体的脂质代谢、能量代谢和免疫过程,导致小鼠肠道微生物失调、肥胖、高脂血症、非酒精性脂肪肝甚至是肝损伤[41-42]. 同时,Chi等还发现,PCBs与高脂饮食同时作用,小鼠肠道F/B比值进一步增加,使得腹部脂肪堆积、肥胖程度进一步增强,这预示着肥胖个体可能更容易受到PCBs暴露的风险[43].
PCBs暴露诱导的肠道微生物组变化除与生物体肥胖相关外,与其他疾病也息息相关. 相关文献证实PCB126暴露后,青蛙肠道中梭杆菌属数量显著增加,其能够产生各种细胞毒素和毒力因子,进一步导致机体肠道炎症的产生[44];同时可导致小鼠盲肠多样性下降,结肠Cyp1a1基因表达上调、肠道炎症标记物干扰素(IFN)、白细胞介素12(IL-12)等增加[45]. 由此说明PCB 126暴露可在一定程度破坏肠道微生物群和宿主代谢,增加了肠道和全身炎症. 肠道微生物群对于宿主而言的一个主要功能是将肝脏产生的初级胆汁酸(BAs)转化为次级胆汁酸,以促进机体进行外源生物转化和能量稳定过程[46-47]. Cheng等研究则发现,小鼠口服PCBs混合物后,其大肠颗粒物或小肠內容物中多个细菌类群丰度与次级胆汁酸之间存在正相关关系,表明PCBs可通过“肠-肝轴”和肠道细菌的共同作用改变宿主胆汁酸的分泌和稳定[48]. 此外,PCBs诱发的遗传和发育毒性也与肠道微生物的组成和结构相关,相关研究发现母体食物中PCBs的暴露会导致幼年小鼠回肠和结肠黏膜屏障出现显著缺陷,而在幼年小鼠中暴露PCBs也会增加肠道炎症和肠道微生物失调的发生[49].
-
PBDEs作为一类溴代阻燃剂被广泛应用于家具、纺织品、电器、电子产品和塑料制品中,但PBDEs是典型的EDCs,能影响个体甲状腺激素的稳态、诱导代谢紊乱、抑制生长等,此外,PBDEs也具有一定的生殖毒性、神经发育毒性、甚至是致畸致癌效应[50]. PBDEs是一类全球性的有机污染物,早在2009年联合国环境规划署正式将四溴联苯醚、五溴联苯醚、六溴联苯醚和七溴联苯醚列入《斯德哥尔摩公约》.
PBDEs可通过动物粪便排出,因此肠道微生物群在该过程中可直接接触污染物[51]. Chen等[52]研究表明成年斑马鱼暴露于环境相关浓度的五溴二苯醚混合物(DE-71)7 d,肠道微生物多样性显著下降且肠道中一些特定的菌属消失(Mycoplasma、Ruminiclostridium、Fusobacterium等),同时包含能量代谢、呼吸、细胞分裂、信号转导和应激反应在内的肠道微生物代谢功能也受到一定程度的影响;Scoville等[53]观察到雄性小鼠的23个肠道细菌类群受到多溴二苯醚的调控;同时,体外实验也表明PBDEs与人体粪便混合时可使使粪便微生物结构产生轻微变化,且能够导致硫代谢、短链脂肪酸以及芳香族物质代谢的紊乱[54]. Li等[55]也证实口服BDE47和BDE99使雄性C57BL/6小鼠肠道细菌多样性减少,但其中与胆汁酸去结合反应相关的Akkermansia muciniphila和Allobaculum spp.数量却有一定的增加;另一项研究也证实BDE47在影响肠道微生物的同时也会在一定程度上使得小鼠肝脏中胆汁酸合成酶Cyp7a1的上调,从而导致粪便和肝脏内胆汁酸分泌量增加[56]. 此外,Wang等[57]还发现,子宫内或哺乳期暴露BDE47会干扰肠道微生物群,导致肠道菌群多样性减少、组成改变和代谢变化,且在高脂饮食组中BDE47还会进一步加重肥胖、肝脏脂肪变性和损伤. 目前,PBDEs对肠道微生物组的影响报道较少,且相关代谢和神经毒性是否是因为肠道微生物变化所引起的也不甚明晰,还需要进一步的研究来揭示PBDEs、肠道微生物及健康效应三者的相互作用.
-
重金属具有蓄积性,在自然环境中难以降解,重金属暴露通常是通过空气、水、食物、土壤等途径发生,并可通过生物富集或食物链等途径引发健康风险, 砷、镉、铅、汞、铜等是常见的重金属污染物. 重金属污染具有多种生物毒性作用,包括肠道微生物群紊乱、致癌作用、氧化应激、DNA损伤、生长发育缓慢、神经和免疫系统损伤等,但是其具体的致病机制尚未阐明[58-60](表2).
-
Cd是一种常见的有毒重金属污染物,具有持久性和生物累积性等特点,广泛应用于工业产品中,如电池、金属镀层、塑料和颜料等,对宿主机体造成严重的健康危害[61]. 越来越多的毒理学实验表明,宿主慢性或急性暴露都会引起肠道菌群的改变和肠道微生态平衡紊乱,从而可能对宿主健康产生不良影响. 此外,Cd暴露能够引起宿主肠道菌群组成和结构的改变且涉及的物种多样(表2),包括人[62]、小鼠[63-65]、蜜蜂[66]、蟾蜍[67]、水生动物斑马鱼[68]、鲤鱼[69-70]、异育银鲫[71]和克氏原螯虾[72]等.
研究表明[63]Cd暴露对不同物种实验动物肠道菌群的影响不同,且对宿主肝脏和肾脏具有毒性作用. 雄性小鼠慢性暴露于低剂量10 mg·L−1的Cd即可显著改变小鼠肠道菌群组成和结构,使厚壁菌门Firmicutes、变形菌门proteobacteria丰度降低,并使得粪便和盲肠中拟杆菌门Bacteroides丰度增加,同时改变肝脏中与糖代谢和脂代谢相关基因的表达,诱导肝脏炎症. 同样地,雄性小鼠Cd 与高脂饮食联合暴露时能够增加肝脏和肾脏中Cd的积累量,加重Cd暴露对小鼠微生物群的影响和对肝、肾功能的损伤[73]. 蜜蜂急性暴露于亚致死浓度Cd中,虽然不影响蜜蜂肠道菌群多样性,但显著该改变蜜蜂肠道微生物组成,使双歧杆菌和乳杆菌丰度降低[66]. 此外,Cd暴露还通过诱导肠道炎症反应、改变肠道屏障功能来破坏肠道稳态[64, 74-76], 导致宿主内毒素血症和炎症反应.
-
Pb是一种典型的高毒性重金属污染物,即使暴露于浓度极低的Pb,也可能引发宿主长期危害. Pb在各种消费品中广泛应用,人和动物可通过空气、水、食物、土壤、油漆、汽油等途径经口摄入、吸入或皮肤吸收暴露于含Pb环境中,引起肠道菌群失调(表2).
长期居住在含Pb等重金属的环境中,能引起人体肠道菌群组成的显著变化,Bacteroides、Roseburia等菌群丰度显著增加,Prevotella 9丰度降低,且在男性和女性群体间存在差异,表现为男性微生物组变化更显著[62]. 在小鼠中,Pb暴露不仅改变肠道微生物组群落结构和多样性,而且极大地影响微生物组的代谢功能,Pb暴露干扰许多代谢途径,包括胆汁酸代谢、氮代谢、能量代谢及诱导氧化应激等[77];Xia等[78]研究表明小鼠长期暴露于0.1 mg·L−1的Pb 15周,盲肠和粪便菌群中厚壁菌门丰度降低,而拟杆菌门在盲肠中下降,在粪便中上升;属水平上,Pb暴露引起Parabacteroides丰度增加,Dehalobacterium丰度降低. 同时代谢组学分析发现Pb暴露进一步引起肝脏代谢变化,影响小鼠氨基酸代谢、三羧酸循环及能量代谢,且肝脏中涉及脂代谢相关的基因表达增加. 成年雄性斑马鱼暴露于10 μg·L−1和30 μg·L−1 Pb中7d,肠道菌群丰富度和多样性发生改变,且厚壁菌门丰度显著增加,变形菌门丰度降低,同时Pb暴露引起斑马鱼肝脏中与糖酵解和脂代谢相关基因表达降低,涉及糖脂代谢、氨基酸代谢及核苷酸代谢等生物学过程发生改变,表明Pb可引起斑马鱼肠道菌群微生态失调和肝脏代谢紊乱[79].
有趣的是,益生菌凝结芽孢杆菌、植物乳杆菌、Faecalibacterium prausnitzii或低聚半乳糖等益生元直接口服或用作膳食补充剂可通过调节肠道菌群来降低体内Pb的积累,减轻因Pb暴露引起的肠道损伤及其相关毒性作用[80-83]. 因此,用益生菌或益生元调节肠道菌群来改善重金属毒性的干预疗法值得进一步研究.
-
As常以砷酸盐的形式存在,广泛分布在水和土壤中. 人体As暴露主要是通过经口摄入被As污染的水和食物等途径. As被认为是人类致癌物,As暴露与肝癌、肺癌、膀胱癌等、糖尿病等代谢性疾病、心血管疾病及骨质疏松和认知功能障碍等相关[84-88].
机体As暴露主要是通过摄入被As污染的水和食物而发生的,因此肠道菌群最易受到影响,As暴露能够改变肠道菌群组成和结构(表2). Dheer等[89]研究发现,雄性小鼠长期暴露于As会引起宿主结肠微生物组组成和结构的改变,并对小鼠氨基酸代谢产生影响. 饮用水中对小鼠进行低剂量10 mg·L-1 As暴露4周,小鼠肠道微生物组成发生改变,同时也会引起肠道微生物组功能及代谢的改变,表现为小鼠肠道中脂质代谢产物、吲哚代谢产物、异黄酮代谢产物和胆汁酸代谢产物的变化[90]. 此外,肠道菌群失调会加剧As引起的毒性损伤,在高脂饮食引起肠道菌群失调的条件下,As暴露导致肝脏和肾脏中As积累量增加,加重As暴露对肝、肾功能的损伤[73]. As暴露不仅引起动物体肠道菌群的改变,人体通过摄入被As污染的水源或食物等,其肠道菌群也会受到影响. Brabec和Hoen等[91-92]研究发现,As暴露破坏了人体肠道菌群,并通过相关性分析发现尿液中As的浓度与肠道菌群丰度呈负相关;Dong等[87]通过比较居住在As暴露区儿童和低As暴露儿童肠道菌群发现,As暴露区儿童粪便菌群中变形菌门丰度增加,且与饮水中As含量呈正相关. 同时,一项研究发现肠道菌群对于保护宿主免受As暴露产生的毒性作用具有重要作用,无菌或经抗生素处理的小鼠粪便中As的浓度减少,相应地器官中的As浓度增加,增加As诱导的毒性作用,表明肠道微生物组破坏影响机体内As的排泄和生物蓄积[93].
-
Hg是一种公认的重金属污染物和食品污染物,广泛应用于化学药品、电子或电器产品中,常温下即可蒸发,Hg蒸气和Hg的化合物多有剧毒. 常以甲基汞的形式存在,Hg及Hg的化合物(甲基汞、氯化汞等)都可经食物链直接或间接在人体内蓄积. 此外,吸入Hg蒸气或裸露的皮肤接触Hg都有可能使Hg进入体内,使机体内细胞、组织和器官等受到损伤.
Hg的微生物转化过程对Hg在地球化学循环中发挥重要作用,微生物既可使Hg甲基化,也可使Hg去甲基化[18]. 目前,已在小鼠、大鼠、鱼体和人体等不同的物种中研究了Hg及Hg的化合物暴露对肠道菌群微生态的影响(表2). 研究表明[94]Hg暴露会影响小鼠的生长发育,导致肠道损伤,诱发小鼠肠道菌群失调和代谢紊乱,并加重小鼠细胞凋亡[94]. 张进[95]等报道,Hg和铜(Cu)联合暴露可使小鼠肠道菌群中有益菌种丰度下降,致病菌丰度显著增加,破坏小鼠肠道正常微生态系统,而肠道菌群的变化又可抑制Hg的排泄,增加Hg在宿主体内的积累量和Hg的毒性作用. 同时发现肠道也是甲基Hg的靶器官之一,甲基Hg能引起小鼠、大鼠和黑头呆鱼(P. promelas)肠道微生物群落结构的改变,并引起肠道神经递质和代谢产物的变化,而肠道神经递质和代谢产物则调节神经元的活性,诱发神经毒性[96-97]. 此外,Hg和Hg的化合物的含量也与肠道菌群及其变化相关. 早期和晚期妊娠期间,母体微生物群组成不同,血液和粪便中总Hg和甲基Hg的浓度有所不同[98]. 另一项研究发现[99]无肠道菌群定植的小鼠,加速了甲基Hg暴露后小鼠肝、肺和脑等组织中Hg的积累[99]. 但是,生物活性物质等营养成分可减轻Hg和Hg的化合物的毒性作用,如补充矿物质元素硒能够使甲基Hg暴露引起的大鼠肠道菌群的变化恢复,并增强甲基Hg的分解和排泄;类黄酮、多酚等植物活性成分和短乳杆菌(Lactobacillus brevis)也可减轻Hg及甲基Hg诱导的毒性作用及其相关的氧化应激和炎症反应,维持肠道稳态[100-101].
-
除了上述有毒重金属外,铬(Cr)、Cu、锰(Mn)等也是环境中常见的重金属污染物,动物机体暴露于这些重金属污染物中,均可能引起机体肠道菌群微生态的改变. Richardson等研究发现大鼠急性暴露于高水平(> 80 mg·kg−1·d−1)铬、钴(Co)和镍(Ni)等重金属中5 d,都能引起肠道变形菌门丰度增加,铬和钴暴露使肠道疣微菌门丰度增加,而镍暴露后疣微菌门丰度下降,同时拟杆菌门中S24-7菌株丰度降低,变形菌门中肠杆菌(Enterobacteriaceae)丰度增加[102]. 小鼠长期暴露于Cr(Ⅵ)能够降低肠道菌群多样性,使厚壁菌门和拟杆菌门丰度下降,Roseburia等产短链脂肪酸菌丰度下降,Verrucomicrobia等丰度增加,加重小鼠氧化应激以及促进小鼠结直肠癌的发生[103]. 另一项在小鼠中的研究发现[104],小鼠经Mn暴露13周后,小鼠肠道微生物多样性降低,但Mn暴露诱导的小鼠肠道微生物群落结构的变化具有性别差异性,雌性小鼠中拟杆菌门和疣微菌门的丰度增加,而在雄性小鼠中表现出丰度降低的趋势;同样地,雌性小鼠中Mn暴露降低了厚壁菌门的丰度,但雄性小鼠中该菌丰度增加. 然而,有趣的是Mn暴露引起的肠道菌群的改变也能够调节体内锰的毒性. 此外,除了在大鼠、小鼠等哺乳动物中的研究,在水生动物中也有相关研究. 鲤鱼暴露于Cr、Cd和Cu的重金属混合物中,发现重金属主要在肾脏和鳃中积累,且暴露导致机体产生氧化应激及肾脏和鳃组织结构的破坏,同时重金属暴露导致鲤鱼肠道微生物群失调,变形菌门、厚壁菌门和梭杆菌门是主要受影响的菌群,短链脂肪酸产生菌和有益菌等丰度下降,而致病菌丰度增加[70, 105].
综上,重金属暴露诱导的机体肠道菌群微生态的改变为研究重金属毒性的机理提供了新的视角,宿主-重金属-微生物群间的相互作用表明,微生物生态失调可能是重金属引起机体健康损害的机制之一. 因此,研究重金属暴露对宿主机体肠道菌群微生态的影响具有重要意义.
-
新污染物是一类新出现的、对环境或人体存在已被证实或潜在危害的化学品的总称,由于缺少完善的管理办法,加之新污染物大多具有环境持久性和生物累积性的特性,在环境介质中有着较强的扩散能力,因此其存在的环境风险不容忽视[113-114]. 通常而言,国际上广泛关注的新污染物主要有四大类,即持久性有机污染物POPs、内分泌干扰物、抗生素和微/纳塑料[115]. 全氟和多氟烷基化合物(PFASs)是一种典型的新型持久性有机污染物POPs. 新烟碱杀虫剂是继有机磷、氨基甲酸酯和拟除虫菊酯之后的新一代农药,越来越多的研究表明其是一种潜在的内分泌干扰物(EDCs). 双酚A及其类似物、三氯生TCS与三氯卡班TCC等均属于EDCs范畴. 研究中广受关注且常见的新污染物大多能够破坏肠道稳态,并由此引发肠、肝、肾损伤、发育障碍、高脂高血糖症等(表3).
-
PFASs是一种典型的新出现的持久性有机污染物,因其具有显著的疏水疏油性、热稳定性、耐腐蚀性、良好的表面活性及抗粘附性等优势,在纺织、化工、电子、医药、农药、航空、电镀材料、家庭用品等多个领域被广泛应用[116]. 目前已在湖泊、大气颗粒物、沉积物中检测到各类PFASs的出现,相关研究也已证实PFASs具有很高的生物蓄积潜力和毒性,且一旦进入生物体内具有较长的半衰期、难以完全生物转化[117-118].
PFASs根据结构不同被分为多个种类,如全氟羧酸(PFCAs)、全氟磺酸(PFSAs)、氟调聚羧酸(FTCAs)、氟调聚磺酸(FTSAs)及氟调聚醇(FTOHs)等[116]. 由于其在生物体内代谢方式及理化特性的不同,不同的PFASs的毒性作用机制和毒性大小也不尽相同. 全氟辛酸(perfluorooctanoic acid,PFOA)暴露后,生物体肝脏作为主要的靶器官极易吸收并累积PFOA,当浓度达到一定水平后便会对肝脏细胞造成严重损伤并导致肝细胞肥大、肝肿大和空泡化[119]. 近年来,“肝-肠轴”的提出为探究污染物肝脏毒性作用机理提供了新的参考,Wang等研究表明急性PFOA暴露导致肠道中Dehalobacterium和拟杆菌(Bacteroides)丰度显著下降;而在慢性暴露过程中,肠道中与修复肝脏损伤相关的乳酸菌和双歧杆菌(Bifidobacterium)等共生菌数量较对照组减少,益生菌丰度的降低会增加肠道通透性,导致急性炎症、肝损伤和糖尿病等[120];此外相关文献也表明肠道微生物组紊乱可导致肠道屏障通透性增加,产生移位的细菌和肠道源性产物的泄漏,最终通过门静脉系统到达肝脏,导致炎症、氧化应激和肝脏疾病的出现[121],而粪菌移植(FMT)治疗可以一定程度上逆转这种菌群与代谢的紊乱[122].
毒性实验证明全氟辛烷磺酸(perfluorooctane sulfonate,PFOS)易在生物体肝脏和血清中累积并与血液胰岛素水平升高、脂代谢异常和神经毒性相关[123]. 暴露于PFOS同样会引起类似变化,相关实验证明小鼠摄食PFOS后与代谢相关的厚壁菌门(Firmicutes)、Bacteroidetes、变形菌门(Proteobacteria)和蓝细菌门(Cyanobacteria)丰度产生一定的变化,进一步会干扰肠道氨基酸(精氨酸、脯氨酸、赖氨酸)、甲烷和短链脂肪酸代谢过程,使脂质和糖代谢发生紊乱,导致宿主肠道屏障功能受损、血糖升高、肝脏脂肪堆积[124-125]. 在离体的小鼠盲肠实验也发现,加入PFOS后会使其中微生物代谢性能显著下降[126].
由于PFOS和PFOA的禁用,ρ-全氟壬苯磺酸钠(OBS)作为替代物应运而生,在物理化学性能方面较PFOS有所增强,但就生物蓄积性和毒性而言却所差无几[127-128]. 2020年的一篇研究表明, 300 μg·L−1 OBS暴露后,斑马鱼肠道中与肝脏代谢物相关的706种菌属相对丰度发生显著变化,因此推测肠道屏障功能的障碍会导致宿主肝脏受损[129]. 此外,全氟丁基磺酸(perfluorobutanesulfonate,PFBS)也是目前常用的一类PFOS替代物,相对于PFOS而言毒性较小但仍具有一定的代谢、视神经和生殖毒性,上述毒性效应已证实与宿主肠道微生失调相关. Chen等[130]研究发现,暴露于PFBS的黑点青鳉(Oryzias melastigma)肠道微生物群的失调是持久和长期存在的,且其后代即便生存在无PFBS的环境,体内的微生物群落也存在明显的紊乱,表明PFBS诱导的宿主微生物群落存在着代际失调. 除上述所提及的几种PFASs外,还在超短链PFASs上有所发现,例如:三氟甲磺酸(TFMS)暴露会导致小鼠盲肠微生物多样性的改变,并进一步引发肝脏炎症反应及脂质代谢损伤[131]. 然而,Chen等[19]研究发现, 益生菌鼠李糖乳杆菌(Lactobacillus rhamnosus)能够抑制PFBS暴露引起的斑马鱼胆固醇积累和胆酸代谢紊乱.
-
新烟碱杀虫剂于上世纪八十年代问世,逐渐作为传统有机磷农药、有机氯农药与拟除虫菊酯杀虫剂的替代品,占据了广泛的杀虫剂市场[132]. 目前常见的新烟碱杀虫剂主要包括吡虫啉(imidacloprid)、啶虫脒(acetamiprid)、噻虫嗪(thiamethoxam)、噻虫啉(thiacloprid)、噻虫胺(clothianidin)、啶虫脒(acetamiprid)、烯啶虫胺(nitenpyram)、呋虫胺(dinotefuran)、氟啶虫酰胺(flonicamid)及硝虫噻嗪(nithiazine)等[132-134]. 多项研究表明,新烟碱杀虫剂存在明显的非靶标生物毒性,暴露于新烟碱农药可能造成蜜蜂等昆虫繁殖能力下降、死亡率升高[135-136],而部分鸟类受新烟碱农药的直接或间接(如食物资源减少等)影响,会出现种群数量的下降[137]与多样性的减少等不良现象[138].
近年来,部分研究关注了新烟碱对哺乳动物的毒性,尤其是通过干扰或破坏机体微生物群落所表现出的毒性. 最近的一项关于小鼠的研究表明,母鼠孕期暴露于烯啶虫胺可导致其雌性后代肠道菌群多样性下降,同时显著改变其菌群结构,包括阿克曼氏菌(Akkermansia)丰度的升高以及Desulfovibrionaceae和乳杆菌丰度的降低[139],损伤肠粘膜,并使食物中的能量被肠道菌群消耗,造成血清甘油三酯、总胆固醇和葡萄糖水平降低[139-140]. 类似的研究将小鼠暴露于呋虫胺、烯啶虫胺和啶虫脒中,显示出氨基酸代谢的紊乱,脂质水平升高,导致氧化应激[141]. 母体暴露于新烟碱农药,其子代表现出的特征与直接暴露于该类农药的毒理学特征存在差异,可能说明母体暴露的情况下,其子代主要受变化后的菌群影响,与直接暴露的毒性效应可能存在不同的作用机理. Yang等[142]对直接暴露后的小鼠肠道菌群进行了研究,雄性小鼠吡虫啉口服暴露造成了肝脏和血清中胆酸水平降低,暴露组的β变形菌、γ变形菌与疣微菌丰度显著增加,Firmicutes/ Bacteroidetes比值(F/B)降低,这种稳态破坏进一步抑制胆酸重吸收,极大地损害了肠屏蔽的完整性,增加机体对有害物质暴露的敏感性. 另外,肠道中的胆汁酸同样可以反作用于肠道菌群,对维持肠道菌群稳态具有重要作用[143],而菌群失衡、肠粘膜损伤可能使机体对外源化学品的防护能力下降,加剧新烟碱农药诱导的机体损伤. Onaru等[144]研究对不同浓度的暴露结果做了补充,总体OTU差别不大的情况下,高浓度的噻虫胺使大鼠F/B值明显减小,放线菌丰度显著升高,导致免疫系统紊乱并抑制幼鼠发育,在低浓度下并无明显表现. 另外,更有研究指出,多种新烟碱杀虫剂同时暴露会造成明显的累积毒性[145],与环境应激产生联合作用[146],造成更为严重和复杂的不良反应.
-
双酚类化合物被广泛应用于生产塑料制品、驱虫剂等[147],常见的有双酚A(BPA)、双酚S(BPS)、双酚F(BPF)及双酚AF(BPAF)与双酚B (BPB)等,报道指出,双酚类化合物可在环境及生物体中被广泛检出,且具有细胞、遗传、生殖与神经毒性[147-148].
口服暴露BPA,能够导致雄鼠肠道菌群的多样性降低,并使变形菌门丰度增加以及阿克曼氏菌的减少,上调了暴露所致的肝脏TNF-α、IL-1β、IL-18和IL-6等炎症细胞因子的表达,造成肠屏蔽功能受损[149-150]. BPA也能够改变菌群代谢,通过研究源自芳香族氨基酸的微生物代谢物的变化,能够反映肠道菌群的表型变化,这种变化使几种与结肠炎发生有拮抗作用的代谢物的水平下降[151],促进肠道炎症的发展. 除口服直接暴露外,母体围产期暴露于BPs同样可能对子代产生不良影响[152-155]. 在围产期暴露于BPs可能会产生跨代效应,如人群队列研究指出,产前暴露于BPA可能会增加儿童患呼吸道疾病的风险[153]. 将孕期小鼠暴露于BPA直至哺乳期结束,观察到亲代肠溶菌酶表达降低,子代结肠通透性增加且炎症易感性增加,在亲代与子代粪便中均发现显著的菌群结构的变化[154-155];但另一方面,雌性子代粪便中双歧杆菌的比例也有所增加[155],可能在一定程度上延缓了疾病的发生. 因此,通过人为接种鼠李糖乳杆菌、双歧杆菌等益生菌,或许可以抑制菌群失衡, 并且Oishi等[156]的研究成果指出接种益生菌会增加BPA的粪便排泄,这说明了益生菌能够强化肠道屏障功能,减少肠道对有害物质的吸收.
此外,暴露于BPS会引起小鼠前额皮层、血清和肠道中神经递质代谢发生紊乱,并可能进而通过脑-肠轴的信号传导造成肠道菌群失衡[157];而斑马鱼暴露于BPS则会引起血管异位萌发,导致早期发育障碍以及全身血液流速降低,提高了其动脉粥样化的风险[158]. Catron等[159]同时评估了多种双酚类化合物对于斑马鱼的暴露风险,在受精后10 d所表现出的发育毒性为BPAF>BPB>BPF≈BPA>BPS,并均对宿主微生物群有破坏作用.
-
三氯生和三氯卡班常用于各类洗护用品以及医用消毒剂中,是一种常用的广谱杀菌剂. 目前在自然水体与土壤等环境介质中均能检测到TCS与TCC残留[160],也有研究指出其在人类血清、尿液、乳汁以及羊水中均存在一定的浓度水平[161-162].
不少研究关注于TCS和TCC对孕妇和胎儿的毒性,包括孕期胎盘、血液传递,以及哺乳期母乳传递,还包括家用产品对婴幼儿的暴露. 有研究指出,由于孕期及哺乳期使用大量含有TCS的洗护用品,导致部分TCS通过母乳向新生儿传递,虽不会降低其肠道菌群多样性,但会造成毛螺菌科(Lachnospiraceae)、韦荣球菌科(Erysipelotrichaceae)与红螺菌科(Rhodospirillaceae)的丰度过高[163],也可能引发菌群衍生代谢物苯基硫酸盐增多,增加其未来发生肾脏损伤的风险[164]. 由于婴幼儿体内微生物群落稳定性较低,生活用品的TCS、TCC暴露极易引起肠道菌群失调,导致变形菌门丰度显著上升,这可能与微生物群抗生素抗性基因的增加有关[1]. 在人粪便菌群体外暴露实验中,高剂量TCS暴露降低了总丰度与物种多样性,并干扰菌群代谢,但停止暴露一段时间后,菌群丰度、多样性与群落结构得到了恢复[165]. 然而,也有大鼠实验指出,生命初期暴露于TCS,对肠道菌群的破坏难以逆转,导致大鼠成年后出现高血糖、血脂积累和肝脏基因表达异常的几率增加,并随着年龄的增长进一步加重,同时也会促进结肠炎症的发展,提高患结肠肿瘤的风险[166]. 同样的,TCC暴露也会导致类似的肝脏损伤,表现为氧化应激增加和代谢紊乱[167],并以肠道微生物为媒介,加重结肠炎症[168].
-
微/纳塑料指的是粒径范围在几微米至几毫米之间(微塑料)或粒径在1 μm以下(纳塑料)的塑料碎片或微粒,这种微小的体积导致其具有独特的迁移富集特征与危害性. 目前,在海洋、淡水及陆地环境中均发现了大量微塑料污染[169],且这种塑料颗粒能够有效地吸附其他污染物,从而对生物体产生联合毒性[170]. 人体通过吸入或者摄食途径暴露于微/纳塑料,可能导致机体局部免疫反应增强,并进一步暴露于微/纳塑料的内源或吸附的污染物[171].
近年的研究表明,微/纳塑料暴露会引起水生生物或哺乳动物肠道菌群的变化. 斑马鱼暴露于微塑料中,导致拟杆菌门和变形菌门丰度显著降低,而厚壁菌门丰度显著增加,属水平上也有29种微生物发生了显著变化,此外还发现了IL-1α、IL-1β和IFN的基因表达水平增加,指示了炎症的发生[172];其他一些研究同样得出了拟杆菌门、变形菌门和厚壁菌门丰度的变化,以及α多样性的下降和胃肠道的损伤[173]. 有趣的是,分别对斑马鱼进行聚酰胺、聚乙烯、聚丙烯、聚氯乙烯和聚苯乙烯不同粒径颗粒的暴露测试,其结果显示肠道损伤和氧化应激是主要健康影响,而这些影响主要与颗粒粒径有关,而与颗粒成分和形状无关[174-175]. 对于小鼠而言,微/纳塑料口服暴露减少了其肠道黏液的分泌,并引起粪便中厚壁菌门与变形菌门丰度的下降,较小粒径暴露引起的属水平上的改变更加明显[176],这种暴露会导致肝脏脂质紊乱、肠屏蔽损伤以及代谢紊乱[177]. 另一项研究指出摄入微/纳塑料增加了小鼠肠道葡萄球菌(Staphylococcus)的丰度而降低了副拟杆菌(Parabacteroides)的丰度,显示出较高的TLR4、AP-1和IRF5表达,以及明显的结肠与十二指肠的炎症反应.
需要注意的是,微/纳塑料通过吸附作用,往往会伴随其他污染物共同暴露,例如邻苯二甲酸酯能够以微塑料作为载体,并产生累积毒性,造成氧化应激、免疫反应、脂质代谢和激素代谢失调等症状[178];类似的,微/纳塑料与TCS共同暴露,会改变TCS在肠道中的分布,加剧其积累,导致肝脏氧化应激加重、代谢紊乱,并产生更明显的神经毒性[179].
-
由于抗生素的广泛使用,加之其在人和动物体内的吸收不完全,大部分摄入的抗生素通过尿液或粪便排出到环境中,往往在各种环境介质中检测到抗生素,如河流和湖泊沉积物、地表水、农业土壤和污水废水等环境中[180-183]. 因此,人和动物可能被动地长期暴露于环境低浓度的抗生素中,其对人和动物的健康不容忽视.
多项研究证实,抗生素引起的微生物群的改变可以在很长一段时间后持续存在. Hagan等[184]研究发现,口服新霉素、万古霉素和甲硝唑可使人体肠道菌群组成和相对丰度发生显著变化,且在停止暴露的180 d后这种差异仍然存在,表明其对微生物组成的影响具有长期作用. 同时抗生素使用后引起炎症信号的增加和树突状细胞的激活,使胆汁酸代谢和色氨酸代谢发生明显改变. 另一项研究同样表明,健康志愿者接受1周的抗生素治疗后,对其肠道菌群的影响持续六个月到两年,包括多样性降低和特定分类群的丧失、抗生素耐药性菌株和抗生素耐药基因的上调[185].
抗生素暴露也可能会通过改变肠道菌群增加某种疾病的严重程度. 艰难梭菌是引起腹泻的主要原因,研究表明单剂量克林霉素或克林霉素和氨苄青霉素两种广谱抗生素联用均能引起小鼠肠道微生物群的显著变化,从而增加对艰难梭菌诱导的结肠炎的易感性,加重腹泻和体重减轻[186-188]. 细胞色素P450单加氧酶(P450)是三大昆虫解毒酶系统之一,抗生素处理诱导蜜蜂肠道微生物菌群失调,进而导致肠道P450解毒酶基因的表达减弱,且微生物菌群失调增加了蜜蜂对噻虫啉的易感性,从而导致蜜蜂存活率降低[188]. 同样地,Coryell等[93]报道当小鼠经头孢哌酮抗生素处理破坏肠道菌群组成后,小鼠组织中重金属As积累量显著增加,相同浓度的As暴露引起小鼠死亡率增加近一半. 然而,也有报道抗生素引起的肠道微生物群的改变对疾病的有益作用. 在狼疮性肾炎经典模型小鼠MRL/lpr中口服氨苄青霉素、新霉素、万古霉素和甲硝唑四联抗生素,能够重塑小鼠肠道菌群组成,增加紧密连接蛋白增强肠道屏障功能,同时通过减少促炎细胞因子IL-17和增加抗炎细胞因子IL-10来改善小鼠炎症程度[189]. 类似的,诺氟沙星不仅减少革兰氏阴性肠道菌群,而且参与IL-10驱动的肠道通透性的调节,改善肠道屏障功能,从而降低自发性细菌性腹膜炎的风险[190].
-
肠道微生物较其他靶器官更为敏感,是能够反映化学品暴露毒性的重要生物标志物. 对三类主要的环境化学品暴露对宿主机体肠道微生态的影响进行了全面综述,并对今后的研究提出了几点展望:
(1)已经发现的化学品暴露通过肠道菌群介导的许多不良影响,包括炎症反应、脂代谢改变、肥胖症、生殖发育和肝肾毒性等,尚缺乏从肠道微生物群的角度探讨化学品暴露影响机体疾病发生发展的机制.
(2)尽管已经建立微生物群紊乱与宿主健康之间的紧密联系,但大多研究仅指出微生物组多样性及菌群组成和结构的改变影响宿主健康,而微生物群是否以及在何种程度上介导了环境化学品暴露的不良健康结局也是亟待解决的科学问题.
(3)仍须确定可以用来敏感地指示肠道微生物群中环境化学品诱导的功能障碍的特定细菌菌株或代谢产物,而非菌群群落. 另一方面,化学品暴露不仅能引起机体肠道微生物群紊乱,而且肠道微生物群相应地也可代谢、降解化学品污染物,但肠道微生物群降解污染物的作用被低估了,这对进一步地研究污染物暴露风险评估也是至关重要的.
(4)微生物群的变化是化学品暴露导致疾病的诱因还是结果仍未有明确的定论,未来的研究应注重化学品暴露诱导的肠道微生物群变化和不良健康结局之间的因果关系,无菌小鼠和体外细胞培养等技术应用促进菌群微生态与健康的相关性研究向因果性转变;
(5)虽然在分析化合物暴露导致的关键微生物和代谢物与健康的关联取得较大进展,但除了找到简单的相关性,化学品暴露-肠道微生态-宿主间相互作用的分子机制有待进一步研究,基因编辑、RNA干扰和过表达等分子生物学技术的应用有助于解析化学品暴露毒性的潜在机制;
(6)最新研究发现肠道菌群通过肠脑轴、肠肝轴、肠肾轴、肠肺轴等远端器官互作来引发机体的不良健康影响,益生菌、益生元等生物活性物质的应用也为改善这些疾病的临床症状提供广阔的应用前景. 因此,为了更好的研究化学品暴露对人群健康的危害作用,应加强人群队列筛查、动物实验和人群验证实验研究. 通过不同人群队列筛查选择某一特定类群中特异性富集或降低的微生物菌种,经分离、培养获得内源性纯菌,以回补方式探讨该菌加重或减轻疾病作用,并探索该菌发挥作用的机制;全基因组测序分析解析该菌基因组中是否含有降解污染物的功能基因,评估实际应用中的降解效能;基因敲除技术应用有助于解析该功能基因的作用,结合动物模型和细胞实验解析其分子机制. 然而,微生物代谢作用也可能通过代谢产生毒性更强的代谢产物而加重污染物预期的毒性,这在今后评估污染物暴露的毒性研究中需要加以考虑.
环境化学品暴露与宿主肠道微生态健康的研究进展与展望
Environmental chemicals exposure and host intestinal microecology: Research progress and prospective
-
摘要: 越来越多的研究证实肠道微生态与宿主机体的健康密切相关. 饮食、环境等外源性因素可显著改变宿主肠道菌群的组成和结构,打破肠道菌群稳态,影响机体代谢从而对宿主健康产生威胁. 人体和动物暴露于各种污染环境时,其肠道微生态是否受到影响,肠道微生态是否为化学品暴露引起健康危害的重要起因,是目前化学品暴露领域的前沿科学问题. 本综述总结了传统持久性有机污染物(POPs)、重金属、新污染物(新型持久性有机污染物、内分泌干扰物、微/纳塑料及抗生素)等三大类主要的环境化学品暴露与宿主肠道微生态稳态的研究进展,讨论了肠道菌群响应环境化学品暴露的变化,及其对宿主健康带来的影响,以期更好地理解环境化学品对人群和动物体的健康风险及其作用机制.
-
关键词:
- 人群暴露 /
- 传统持久性有机污染物 /
- 重金属 /
- 新污染物 /
- 肠道微生态.
Abstract: A growing studies have confirmed that the gut microbiota was closely related to the host health. Exogenous factors such as diet and environmental exposure can significantly alter the composition and structure of host intestinal flora, break the homeostasis of intestinal flora, affect the metabolism of host and threaten host health. Whether the intestinal microecology of human beings and animals is affected when they are exposed to various polluted environments and whether the intestinal microecology is an important factor of health risks caused by environmental chemical exposure are the frontier scientific issues in the field of chemical exposure. The current review summarized the research progress on the associations between three major environmental chemicals and host intestinal microecology, including legacy persistent organic pollutants (POPs), heavy metals and emerging pollutants (emerging POPs, endocrine disrupting chemicals, micro-/nanoplastics and antibiotics). Then we discussed the changes of intestinal flora in response to environmental chemical exposure and the adversely effects on host health. It can provide new sight and evidence on the health risks and related mechanisms of environmental chemicals exposure among human and animals. -

-
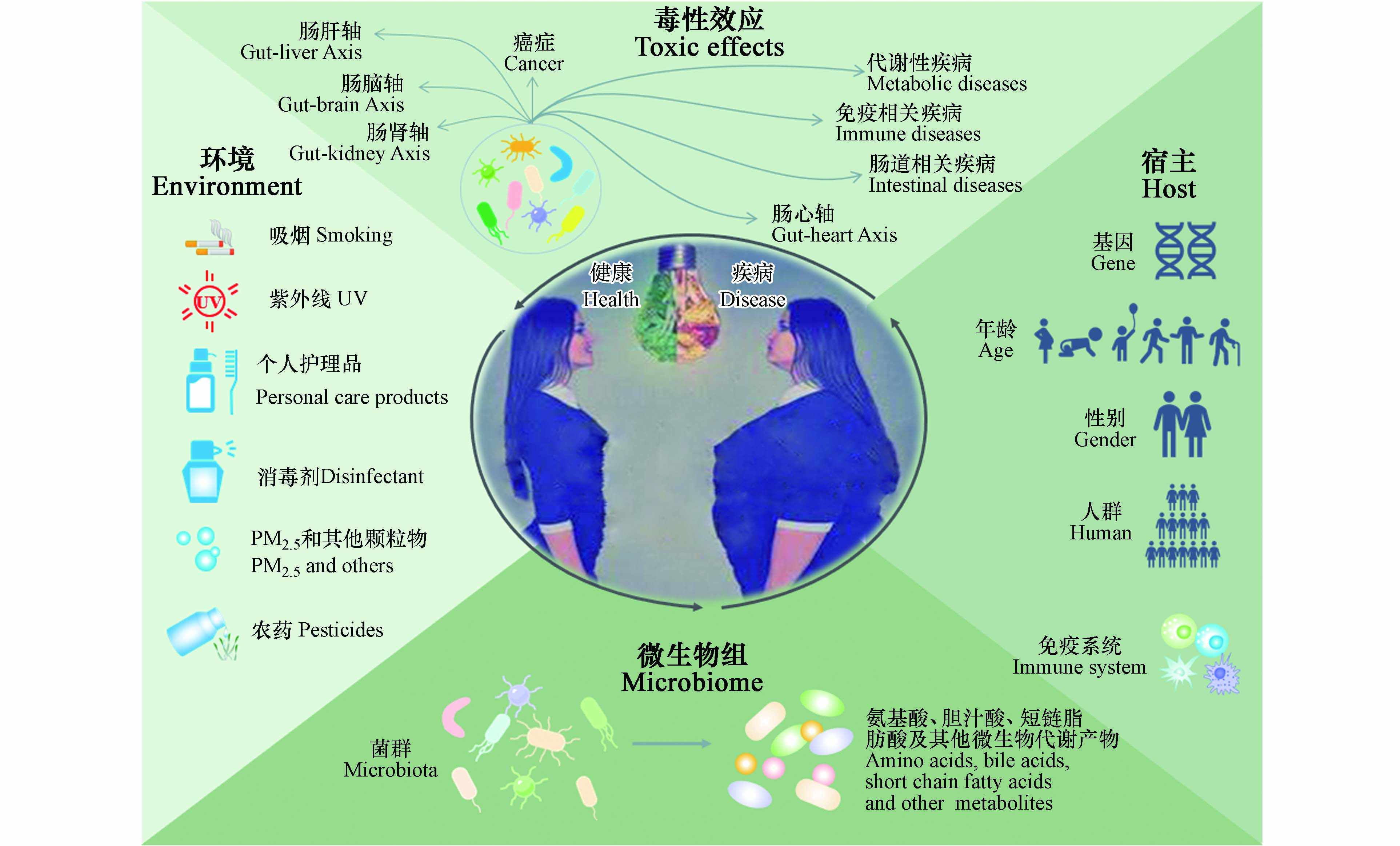
图 1 环境暴露、肠道微生物群和宿主之间的相互作用
Figure 1. The associations among human exposure to environmental pollutants, gut microbiota and host
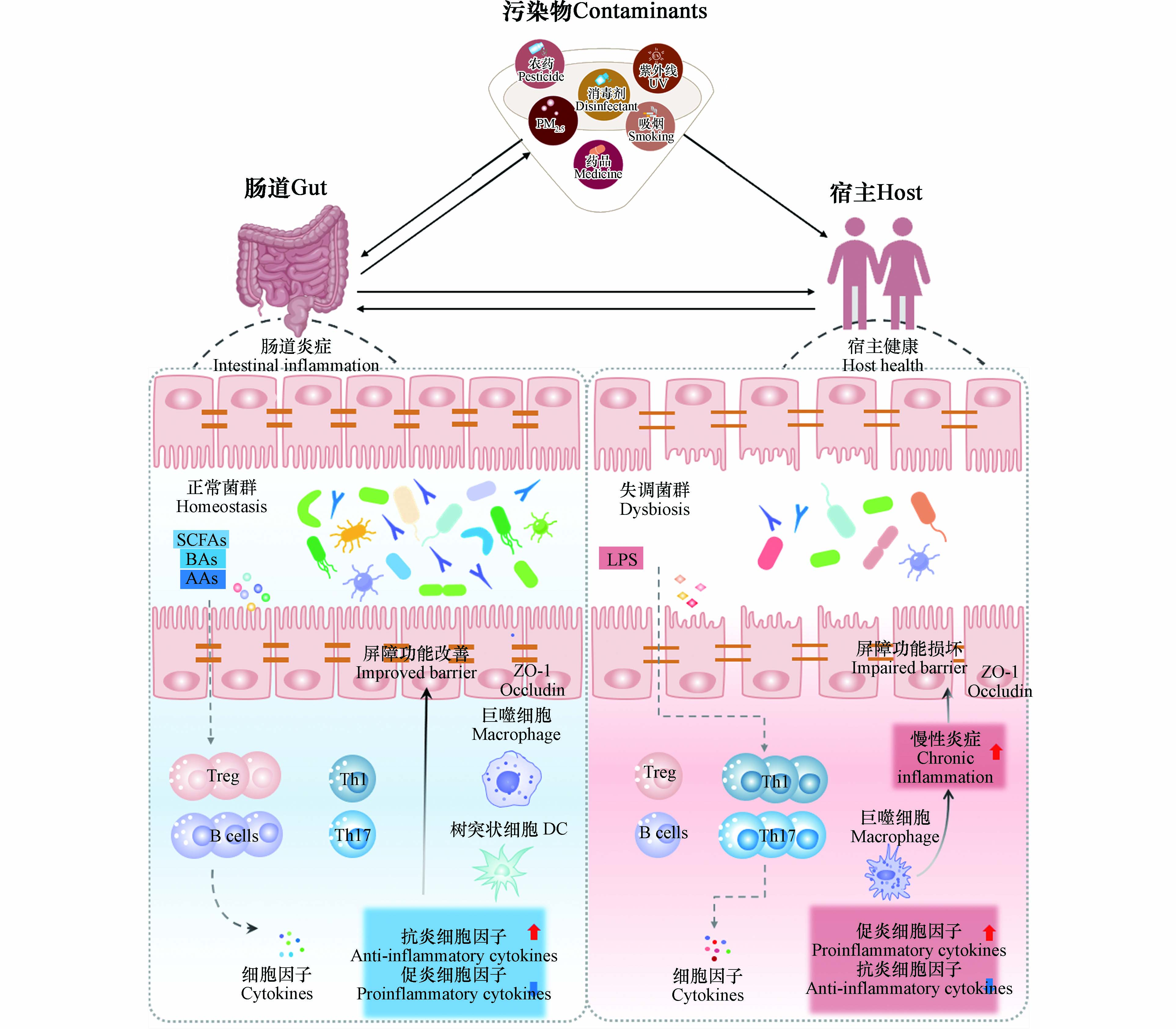
图 2 肠道微生物群介导的环境污染物毒性机制
Figure 2. Mechanism of gut microbiota-mediated toxicity of environmental contaminants
表 1 持久性有机污染物POPs暴露驱动的宿主肠道微生物群组成变化
Table 1. POPs-driven changes in gut microbiome composition in host
污染物
Contaminants宿主
Host剂量
Dose暴露时间/方式
Exposure time/modes菌群变化
Changes毒性效应
Effects参考文献References PAHs 蠕虫 3%土壤干重 21 d/土壤 Alicyclobacillus, Flavobacterium↑; Acinetobacter, Cupriavidus↓ 存活率、繁殖率下降 [28] PAHs 人 — 工厂混合区 Actinobacteria↑; Bacillus↓ 儿童哮喘风险 [30] PAHs 尖齿胡鲶 70 mg·kg−1 3 d/肌肉注射 Pseudomonas aeruginosa↑ — [32] PAHs 小鼠 50 mg·kg−1 28 d/口服灌胃 Bacteroides, Desulfovibrionaceae↑; Lactobacillus, Akkermansia↓ 肠道组织病变 [37] PCBs 小鼠 50 μg·kg−1 6 w/口服灌胃 Helicobacter↑; Allobaculum↓ 肥胖、高脂血症、脂肪肝 [41] PCBs 青蛙 7.3 ng·g−1 85 d/喂食 Pseudomonas↑; Aminobacter↓ 肠道炎症 [44] PCBs 小鼠 6 mg·kg−1 产前2 w产后3 w/喂食 Proteobacteria↑; Alistipes, S24-7↓ 肠道炎症、遗传毒性 [49] PBDEs 斑马鱼 5.0 ng·L−1 7 d/养殖水 Mycoplasma, Ruminiclostridium, Fusobacterium↓ 肠道炎症、氧化应激 [52] PBDEs 小鼠 0.2 mg·kg−1 产后21 d/口服灌胃 S24-7↑; Ruminococcaceae, Rikenellaceae↓ 胆酸代谢失调 [56] PBDEs 小鼠 0.2 mg·kg−1 14 w/口服灌胃 Dorea, Lactococcus↑; Turicibacter, Anaerotruncus↓ 肝脏脂肪变性、葡萄糖稳态紊乱 [57] 注:↑-增加;↓-降低. Note: ↑-increase; ↓-decrease  下载: 导出CSV
下载: 导出CSV
表 2 重金属暴露驱动的宿主肠道微生物群组成变化
Table 2. Heavy metal-driven changes in gut microbiome composition in host
污染物
Contaminants宿主
Host剂量
Dose暴露时间/方式
Exposure time/modes菌群变化
Changes毒性效应
Effects参考文献
References
Cd人 — 铅锌矿冶炼区 Lachnospiraceae, Bacteroides, Roseburia↑;
Prevotella 9↓— [62] 小鼠 100 mg·L−1 8 w/饮水 Alistipes, Odoribacter↑
Mollicutes, Ruminococcaceae, Akkermansia↓— [65] 斑马鱼
胚胎5 μg·L−1 7 d/无菌培养液 Proteobacteria↑; Firmicutes↓ 神经发育毒性,降低存活率 [68] Pb 小鼠 100 mg·L−1 10 w/饮水 Desulfovibrio, Prevotella, Roseburia↑ 肝肾损伤 [73] 鲤鱼 1 mg·L−1 42 d/养殖水 Bacteroidetes, Fusobacteria, Plesiomonas↑; Actinobacteria; Akkermansia↓ 肠道组织病变 [106] As 小鼠 100 mg·L−1 10 w/饮水 Bacteroides, Butyricimonas, Dorea, Lactobacillus↑ 肝肾损伤 [73] 大鼠 50 mg·L−1 21 d/饮水 Bacteroidetes, Epsilonbacteraeota↑;
Firmicutes↓神经发育毒性 [107] 斑马鱼 100 μg·L−1 20 d/养殖水 Acinetobacter, Sediminibacterium, Methylotenera↑; Bdellovibrio, Pseudomonas↓ — [108] 蚯蚓 40 mg·L−1 28 d/土壤 Proteobacteria, Enterobacteriaceae↑; Acidobacteria, Chloroflexi↓ — [109] Hg 大鼠 0.4 μg·mL−1 24 h/口服 Frmicutes↑; Bacteroidetes, Proteobacteria↓; Lactobacillaceae, Bacteroidaceae, Streptococcaceae, Sutterellaceae↓;
Desulfovibrionaceae, Helicobacteraceae, Peptococcaceae, Rhodospirillaceae↑神经毒性 [95] 小鼠 160 mg·L−1 3 d/饮水 Oscillospira, Desulfovibrio↑; S24-7, Acinetobacter↓ 体重减轻、氧化应激 [110] 鸡 250 mg·L−1 90 d/饮水 Proteobacteria, Tenericutes↑; Spirochaetes↓ 体重减轻、肠道损伤 [111] 斑马鱼 10 ng·mL−1 14 d/养殖水 Aeromonas, Cetobacterium↓ 游动受损 [112] 注:↑-增加;↓-降低. Note: ↑-increase; ↓-decrease
下载: 导出CSV
表 3 新污染物暴露驱动的宿主肠道微生物群组成变化
Table 3. Emerging contaminants-driven changes in gut microbiome composition in host
污染物
Contaminants宿主
Host剂量
Dose暴露时间/方式
Exposure time/modes菌群变化
Changes毒性效应
Effects参考文献References PFAS PFOA 小鼠 30 mg·kg−1 14 d/口服灌胃 Dorea↑; Lactobacillus, Bifidobacterium↓ 减轻体重、肝损伤 [120] PFOS 小鼠 3 mg·kg−1 7 w/喂食 Turicibacter↑; Bacteroides↓ 脂质和糖代谢紊乱 [124] OBS 斑马鱼 300 μg·L−1 21 d/养殖水 Fusobacteria, Verrucomicrobia↑; Verrucomicrobia, Proteobacteria↓ 肝损伤、肠屏障
受损[129] PFBS 斑马鱼 100 μg·L−1 28 d/养殖水 Roseburia, Faecalibacterium↑; Cetobacterium↓ 胆固醇积累、胆酸代谢紊乱 [19] EDCs NEOs 小鼠 4 mg·kg−1 孕鼠暴露后后代11 w/口服灌胃 Prevotella↑; Desulfovibrionaceae, Lactobacillus↓ 肠道损伤、脂质代谢紊乱 [141] NEOs 大鼠 300 mg·kg−1 28 d/口服灌胃 Bifidobacterium↑; Lachnospiraceae, F/B↓ 免疫系统紊乱、抑制幼鼠发育 [144] BPA 小鼠 50 μg·kg−1 24 w/喂食 Proteobacteria↑; Verrucomicrobia, Akkermansia↓ 肝纤维化、肠屏障功能受损 [150] BPA 小鼠 50 mg·kg−1 孕期至断奶整个
期间/饮水Desulfovibrio, Lactococcus, Allobaculum↑; Sutterella↓ 肠道通透性和炎症易感性增加 [155] BPS 斑马鱼胚胎 100 μg·L−1 16 d/养殖水 — 血管异位萌发、早期发育障碍 [158] BPs 斑马鱼胚胎 45 μM·L−1 10 d/养殖水 Rheinheimera, Pseudomonas↑; Chitinimonas↓ 神经发育毒性 [159] TCS 人 — 母乳 Lachnospiraceae, Erysipelotrichaceae, Rhodospirillaceae↑ 增加新生儿肾脏损伤风险 [163] TCS 大鼠 50 mg·kg−1 3 w/口服灌胃 Bacteroidetes↑;Verrucomicrobia, Akkermansia↓ 高血糖、高血脂症 [167] TCC 小鼠 80 mg·L−1 3 w/饮水 Proteobacteria↑; Bifidobacterium, Akkermansia↓ 代谢紊乱、促进结肠肿瘤产生 [169] MPs MPs 斑马鱼 1000 μg·L−1 14 d/养殖水 Firmicutes↑; Bacteroidetes, Proteobacteria↓ 肠道粘液增加、炎症损伤 [173] MPs 小鼠 1000 μg·L−1 5 w/饮水 Firmicutes, α-Proteobacteria↓ 肝脂质代谢紊乱、肠道损伤 [177] 抗生素 抗生素 人 500 mg·d−1 5 d/口服 Enterobacteriaceae↑; Ruminococcaceae↓ 免疫功能紊乱 [185] 抗生素 小鼠 200 μg·d−1 腹腔注射 Enterobacteriaceae↑; Lachnospiraceae, Barnesiella↓ 增加结肠炎易感性、加重腹泻 [187-188] 抗生素 小鼠 — 饮水 Lactobacillales↑; E. faecalis, E. coli↓ 改善肠道通透性、减轻炎症 [189-190] 注:↑-增加;↓-降低. Note: ↑-increase; ↓-decrease
下载: 导出CSV
-
[1] RIBADO J V, LEY C, HAGGERTY T D, et al. Household triclosan and triclocarban effects on the infant and maternal microbiome [J]. EMBO Molecular Medicine, 2017, 9(12): 1732-1741. doi: 10.15252/emmm.201707882 [2] LUKOWICZ C, ELLERO-SIMATOS S, RÉGNIER M, et al. Metabolic effects of a chronic dietary exposure to a low-dose pesticide cocktail in mice: Sexual dimorphism and role of the constitutive androstane receptor [J]. Environmental Health Perspectives, 2018, 126(6): 067007. doi: 10.1289/EHP2877 [3] YAN X Y, CHEN X S, TIAN X L, et al. Co-exposure to inorganic arsenic and fluoride prominently disrupts gut microbiota equilibrium and induces adverse cardiovascular effects in offspring rats [J]. The Science of the Total Environment, 2021, 767: 144924. doi: 10.1016/j.scitotenv.2020.144924 [4] WANG W J, XIE Z T, LIN Y, et al. Association of inorganic arsenic exposure with type 2 diabetes mellitus: A meta-analysis [J]. Journal of Epidemiology and Community Health, 2014, 68(2): 176-184. doi: 10.1136/jech-2013-203114 [5] ZHANG Q, HAO L C, HONG Y. Exploring the multilevel effects of triclosan from development, reproduction to behavior using Drosophila melanogaster [J]. The Science of the Total Environment, 2021, 762: 144170. doi: 10.1016/j.scitotenv.2020.144170 [6] REGNAULT C, USAL M, VEYRENC S, et al. Unexpected metabolic disorders induced by endocrine disruptors in Xenopus tropicalis provide new lead for understanding amphibian decline [J]. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(19): E4416-E4425. [7] ADAMOVSKY O, BUERGER A N, VESPALCOVA H, et al. Evaluation of microbiome-host relationships in the zebrafish gastrointestinal system reveals adaptive immunity is a target of bis(2-ethylhexyl) phthalate (DEHP) exposure [J]. Environmental Science & Technology, 2020, 54(9): 5719-5728. [8] YUEH M F, HE F, CHEN C, et al. Triclosan leads to dysregulation of the metabolic regulator FGF21 exacerbating high fat diet-induced nonalcoholic fatty liver disease [J]. Proceedings of the National Academy of Sciences of the United States of America, 2020, 117(49): 31259-31266. doi: 10.1073/pnas.2017129117 [9] CUI H Y, CHANG Y Q, JIANG X F, et al. Triphenyl phosphate exposure induces kidney structural damage and gut microbiota disorders in mice under different diets [J]. Environment International, 2020, 144: 106054. doi: 10.1016/j.envint.2020.106054 [10] HU J Z, RAIKHEL V, GOPALAKRISHNAN K, et al. Effect of postnatal low-dose exposure to environmental chemicals on the gut microbiome in a rodent model [J]. Microbiome, 2016, 4(1): 26. doi: 10.1186/s40168-016-0173-2 [11] LIU Y J, QIAO N H, DIAO Q Y. et al. Thiacloprid exposure perturbs the gut microbiota and reduces the survival status in honeybees [J]. Journal of Hazardous Materials, 2020, 389: 121818. doi: 10.1016/j.jhazmat.2019.121818 [12] SCHMIDT T S B, RAES J, BORK P. The human gut microbiome: From association to modulation [J]. Cell, 2018, 172(6): 1198-1215. doi: 10.1016/j.cell.2018.02.044 [13] YATSUNENKO T, REY F E, MANARY M J, et al. Human gut microbiome viewed across age and geography [J]. Nature, 2012, 486(7402): 222-227. doi: 10.1038/nature11053 [14] GOODRICH J K, WATERS J L, POOLE A C, et al. Human genetics shape the gut microbiome [J]. Cell, 2014, 159(4): 789-799. doi: 10.1016/j.cell.2014.09.053 [15] BARTON W, PENNEY N C, CRONIN O, et al. The microbiome of professional athletes differs from that of more sedentary subjects in composition and particularly at the functional metabolic level [J]. Gut, 2018, 67(4): 625-633. [16] DUBINKINA V B, TYAKHT A V, ODINTSOVA V Y, et al. Links of gut microbiota composition with alcohol dependence syndrome and alcoholic liver disease [J]. Microbiome, 2017, 5(1): 1-14. doi: 10.1186/s40168-016-0209-7 [17] YOUNAN S, SAKITA G Z, ALBUQUERQUE T R, et al. Chromium(VI) bioremediation by probiotics [J]. Journal of the Science of Food and Agriculture, 2016, 96(12): 3977-3982. doi: 10.1002/jsfa.7725 [18] YANG T T, LIU Y, TAN S, et al. The role of intestinal microbiota of the marine fish (Acanthopagrus latus) in mercury biotransformation [J]. Environmental Pollution (Barking, Essex:1987), 2021, 277: 116768. doi: 10.1016/j.envpol.2021.116768 [19] CHEN L G, LAM J C W, TANG L Z, et al. Probiotic modulation of lipid metabolism disorders caused by perfluorobutanesulfonate pollution in zebrafish [J]. Environmental Science & Technology, 2020, 54(12): 7494-7503. [20] LOHMANN R, BREIVIK K, DACHS J, et al. Global fate of POPs: Current and future research directions [J]. Environmental Pollution (Barking, Essex:1987), 2007, 150(1): 150-165. doi: 10.1016/j.envpol.2007.06.051 [21] METIDJI A, OMENETTI S, CROTTA S, et al. The environmental sensor AHR protects from inflammatory damage by maintaining intestinal stem cell homeostasis and barrier integrity [J]. Immunity, 2018, 49(2): 353-362.e5. doi: 10.1016/j.immuni.2018.07.010 [22] PÉREZ T, BALCÁZAR J L, RUIZ-ZARZUELA I, et al. Host-microbiota interactions within the fish intestinal ecosystem [J]. Mucosal Immunology, 2010, 3(4): 355-360. doi: 10.1038/mi.2010.12 [23] CERNIGLIA C E. Biodegradation of polycyclic aromatic hydrocarbons [J]. Biodegradation, 1992, 3(2): 351-368. [24] DOUBEN P E T. PAHs: An Ecotoxicological Perspective [M]. Chichester, UK: John Wiley & Sons, Ltd, 2003. [25] HICKEN C E, LINBO T L, BALDWIN D H, et al. Sublethal exposure to crude oil during embryonic development alters cardiac morphology and reduces aerobic capacity in adult fish [J]. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(17): 7086-7090. doi: 10.1073/pnas.1019031108 [26] DEBOFSKY A, XIE Y W, JARDINE T D, et al. Effects of the husky oil spill on gut microbiota of native fishes in the North Saskatchewan River, Canada [J]. Aquatic Toxicology, 2020, 229: 105658. doi: 10.1016/j.aquatox.2020.105658 [27] BAYHA K M, ORTELL N, RYAN C N, et al. Crude oil impairs immune function and increases susceptibility to pathogenic bacteria in southern flounder [J]. PLoS One, 2017, 12(5): e0176559. doi: 10.1371/journal.pone.0176559 [28] DING J, ZHU D, WANG H T, et al. Dysbiosis in the gut microbiota of soil fauna explains the toxicity of tire tread particles [J]. Environmental Science & Technology, 2020, 54(12): 7450-7460. [29] FOULADI F, BAILEY M J, PATTERSON W B, et al. Air pollution exposure is associated with the gut microbiome as revealed by shotgun metagenomic sequencing [J]. Environment International, 2020, 138: 105604. doi: 10.1016/j.envint.2020.105604 [30] HU J Y, BAO Y L, ZHU Y Q, et al. The preliminary study on the association between PAHs and air pollutants and microbiota diversity[J]. Archives of Environmental Contamination and Toxicology, 2020, 79(3): 321-332 [31] VARI H K, ROSLUND M I, OIKARINEN S, et al. Associations between land cover categories, gaseous PAH levels in ambient air and endocrine signaling predicted from gut bacterial metagenome of the elderly [J]. Chemosphere, 2021, 265: 128965. [32] KARAMI A, CHRISTIANUS A, ISHAK Z, et al. Use of intestinal Pseudomonas aeruginosa in fish to detect the environmental pollutant benzo[a]pyrene [J]. Journal of Hazardous Materials, 2012, 215/216: 108-114. doi: 10.1016/j.jhazmat.2012.02.038 [33] DeBOFSKY A, XIE Y W, GRIMARD C, et al. Differential responses of gut microbiota of male and female fathead minnow (Pimephales promelas) to a short-term environmentally-relevant, aqueous exposure to benzo[a]pyrene [J]. Chemosphere, 2020, 252: 126461. doi: 10.1016/j.chemosphere.2020.126461 [34] ZHAO Y, LIU H, WANG Q, et al. The effects of benzo[a]pyrene on the composition of gut microbiota and the gut health of the juvenile sea cucumber Apostichopus japonicus Selenka [J]. Fish & Shellfish Immunology, 2019, 93: 369-379. [35] XIE S L, ZHOU A G, XU N, et al. Benzo[a]pyrene induces microbiome dysbiosis and inflammation in the intestinal tracts of western mosquitofish (Gambusia affinis) and zebrafish (Danio rerio) [J]. Fish & Shellfish Immunology, 2020, 105: 24-34. [36] DeBOFSKY A, XIE Y W, CHALLIS J K, et al. Responses of juvenile fathead minnow (Pimephales promelas) gut microbiome to a chronic dietary exposure of benzo[a]pyrene [J]. Environmental Pollution (Barking, Essex:1987), 2021, 278: 116821. doi: 10.1016/j.envpol.2021.116821 [37] RIBIÈRE C, PEYRET P, PARISOT N, et al. Oral exposure to environmental pollutant benzo[a]pyrene impacts the intestinal epithelium and induces gut microbial shifts in murine model [J]. Scientific Reports, 2016, 6: 31027. doi: 10.1038/srep31027 [38] QUINETE N, SCHETTGEN T, BERTRAM J, et al. Occurrence and distribution of PCB metabolites in blood and their potential health effects in humans: A review [J]. Environmental Science and Pollution Research, 2014, 21(20): 11951-11972. doi: 10.1007/s11356-014-3136-9 [39] TANG-PÉRONARD J L, ANDERSEN H R, JENSEN T K, et al. Endocrine-disrupting chemicals and obesity development in humans: A review [J]. Obesity Reviews:an Official Journal of the International Association for the Study of Obesity, 2011, 12(8): 622-636. doi: 10.1111/j.1467-789X.2011.00871.x [40] CHOI J J, EUM S Y, RAMPERSAUD E, et al. Exercise attenuates PCB-induced changes in the mouse gut microbiome [J]. Environmental Health Perspectives, 2013, 121(6): 725-730. doi: 10.1289/ehp.1306534 [41] CHI Y L, WANG H O, LIN Y, et al. Gut microbiota characterization and lipid metabolism disorder found in PCB77-treated female mice [J]. Toxicology, 2019, 420: 11-20. doi: 10.1016/j.tox.2019.03.011 [42] CHI Y L, LIN Y, LU Y Y, et al. Gut microbiota dysbiosis correlates with a low-dose PCB126-induced dyslipidemia and non-alcoholic fatty liver disease [J]. Science of the Total Environment, 2019, 653: 274-282. doi: 10.1016/j.scitotenv.2018.10.387 [43] CHI Y L, LIN Y, ZHU H M, et al. PCBs-high-fat diet interactions as mediators of gut microbiota dysbiosis and abdominal fat accumulation in female mice [J]. Environmental Pollution, 2018, 239: 332-341. doi: 10.1016/j.envpol.2018.04.001 [44] KOHL K D, CARY T L, KARASOV W H, et al. Larval exposure to polychlorinated biphenyl 126 (PCB-126) causes persistent alteration of the amphibian gut microbiota [J]. Environmental Toxicology and Chemistry, 2015, 34(5): 1113-1118. doi: 10.1002/etc.2905 [45] PETRIELLO M C, HOFFMAN J B, VSEVOLOZHSKAYA O, et al. Dioxin-like PCB 126 increases intestinal inflammation and disrupts gut microbiota and metabolic homeostasis [J]. Environmental Pollution (Barking, Essex: 1987), 2018, 242(Pt A): 1022-1032. [46] SPANOGIANNOPOULOS P, BESS E N, CARMODY R N, et al. The microbial pharmacists within us: A metagenomic view of xenobiotic metabolism [J]. Nature Reviews Microbiology, 2016, 14(5): 273-287. doi: 10.1038/nrmicro.2016.17 [47] CARMODY R N, TURNBAUGH P J. Host-microbial interactions in the metabolism of therapeutic and diet-derived xenobiotics [J]. The Journal of Clinical Investigation, 2014, 124(10): 4173-4181. doi: 10.1172/JCI72335 [48] CHENG S L, LI X S, LEHMLER H J, et al. Gut microbiota modulates interactions between polychlorinated biphenyls and bile acid homeostasis [J]. Toxicological Sciences, 2018, 166(2): 269-287. [49] RUDE K M, PUSCEDDU M M, KEOGH C E, et al. Developmental exposure to polychlorinated biphenyls (PCBs) in the maternal diet causes host-microbe defects in weanling offspring mice [J]. Environmental Pollution, 2019, 253: 708-721. doi: 10.1016/j.envpol.2019.07.066 [50] MEEKER J D, JOHNSON P I, CAMANN D, et al. Polybrominated diphenyl ether (PBDE) concentrations in house dust are related to hormone levels in men [J]. Science of the Total Environment, 2009, 407(10): 3425-3429. doi: 10.1016/j.scitotenv.2009.01.030 [51] HUWE J K, SMITH D J. Accumulation, whole-body depletion, and debromination of decabromodiphenyl ether in male sprague-dawley rats following dietary exposure [J]. Environmental Science & Technology, 2007, 41(7): 2371-2377. [52] CHEN L G, HU C Y, LOK-SHUN L N, et al. Acute exposure to PBDEs at an environmentally realistic concentration causes abrupt changes in the gut microbiota and host health of zebrafish [J]. Environmental Pollution, 2018, 240: 17-26. doi: 10.1016/j.envpol.2018.04.062 [53] SCOVILLE D K, LI C Y, WANG D F, et al. Polybrominated diphenyl ethers and gut microbiome modulate metabolic syndrome-related aqueous metabolites in mice [J]. Drug Metabolism and Disposition:the Biological Fate of Chemicals, 2019, 47(8): 928-940. doi: 10.1124/dmd.119.086538 [54] CRUZ R, PALMEIRA J D, MARTINS Z E, et al. Multidisciplinary approach to determine the effect of polybrominated diphenyl ethers on gut microbiota [J]. Environmental Pollution, 2020, 260: 113920. doi: 10.1016/j.envpol.2020.113920 [55] LI X M, DONG S J, ZHANG W, et al. Global occurrence of polybrominated diphenyl ethers and their hydroxylated and methoxylated structural analogues in an important animal feed (fishmeal) [J]. Environmental Pollution, 2018, 234: 620-629. doi: 10.1016/j.envpol.2017.11.059 [56] GOMEZ M V, DUTTA M, SUVOROV A, et al: Early life exposure to environmental contaminants (BDE-47, TBBPA, and BPS) produced persistent alterations in fecal microbiome in adult male mice [J]. Toxicological Sciences, 2021, 179(1): 14-30. [57] WANG D Z, YAN J, TENG M M, et al. In utero and lactational exposure to BDE-47 promotes obesity development in mouse offspring fed a high-fat diet: Impaired lipid metabolism and intestinal dysbiosis [J]. Archives of Toxicology, 2018, 92(5): 1847-1860. doi: 10.1007/s00204-018-2177-0 [58] RAJESWARI M, RAMYA S. Assessment of heavy metals in aallus and their impacts on human [J]. International Journal of Scientific and Research Publications, 2014, 4: 275308342. [59] IDREES M, BATOOL S. Environmental risk assessment of chronic arsenic in drinking water and prevalence of type-2 diabetes mellitus in Pakistan [J]. Environmental Technology, 2020, 41(2): 232-237. doi: 10.1080/09593330.2018.1494754 [60] JIN Y X, LIU L, ZHANG S B, et al. Chromium alters lipopolysaccharide-induced inflammatory responses both in vivo and in vitro [J]. Chemosphere, 2016, 148: 436-443. doi: 10.1016/j.chemosphere.2016.01.057 [61] ALGHASHAM A, SALEM T A, MEKI A R M. Effect of cadmium-polluted water on plasma levels of tumor necrosis factor-α, interleukin-6 and oxidative status biomarkers in rats: Protective effect of curcumin [J]. Food and Chemical Toxicology:an International Journal Published for the British Industrial Biological Research Association, 2013, 59: 160-164. doi: 10.1016/j.fct.2013.05.059 [62] SHAO M M, ZHU Y. Long-term metal exposure changes gut microbiota of residents surrounding a mining and smelting area [J]. Scientific Reports, 2020, 10: 4453. doi: 10.1038/s41598-020-61143-7 [63] ZHANG S B, JIN Y X, ZENG Z Y, et al. Subchronic exposure of mice to cadmium perturbs their hepatic energy metabolism and gut microbiome [J]. Chemical Research in Toxicology, 2015, 28(10): 2000-2009. doi: 10.1021/acs.chemrestox.5b00237 [64] HE X W, Qi Z D, Hou H, et al. Structural and functional alterations of gut microbiome in mice induced by chronic cadmium exposure [J]. Chemosphere, 2020, 246: 125747. doi: 10.1016/j.chemosphere.2019.125747 [65] ZHAI Q X, LI T Q, YU L L, et al. Effects of subchronic oral toxic metal exposure on the intestinal microbiota of mice [J]. Science Bulletin, 2017, 62(12): 831-840. doi: 10.1016/j.scib.2017.01.031 [66] ROTHMAN J A, LEGER L, KIRKWOOD J S, et al. Cadmium and selenate exposure affects the honey bee microbiome and metabolome, and bee-associated bacteria show potential for bioaccumulation [J]. Applied and Environmental Microbiology, 2019, 85(21): e01411-e01419. [67] KIMURA I, MIYAMOTO J, OHUE-KITANO R, et al. Maternal gut microbiota in pregnancy influences offspring metabolic phenotype in mice [J]. Science, 2020, 367(6481): eaaw8429. doi: 10.1126/science.aaw8429 [68] XIA Y, ZHU J W, XU Y J, et al. Effects of ecologically relevant concentrations of cadmium on locomotor activity and microbiota in zebrafish [J]. Chemosphere, 2020, 257: 127220. doi: 10.1016/j.chemosphere.2020.127220 [69] CHANG X L, Li H, Feng J C, et al. Effects of cadmium exposure on the composition and diversity of the intestinal microbial community of common carp (Cyprinus carpio L. ) [J]. Ecotoxicology and Environmental Safety, 2019, 171: 92-98. doi: 10.1016/j.ecoenv.2018.12.066 [70] KAKADE A, SALAMA E S, FENG P Y, et al. Long-term exposure of high concentration heavy metals induced toxicity, fatality, and gut microbial dysbiosis in common carp, Cyprinus carpio [J]. Environmental Pollution, 2020, 266: 115293. doi: 10.1016/j.envpol.2020.115293 [71] WANG N, GUO Z Y, ZHANG Y L, et al. Effect on intestinal microbiota, bioaccumulation, and oxidative stress of Carassius auratus gibelio under waterborne cadmium exposure [J]. Fish Physiology and Biochemistry, 2020, 46(6): 2299-2309. doi: 10.1007/s10695-020-00870-0 [72] ZHANG Y, Li Z Y, KHOLODKEVICH S, et al. Effects of cadmium on intestinal histology and microbiota in freshwater crayfish (Procambarus clarkii) [J]. Chemosphere, 2020, 242: 125105. doi: 10.1016/j.chemosphere.2019.125105 [73] LIU T, LIANG X, LEI C, et al. High-fat diet affects heavy metal accumulation and toxicity to mice liver and kidney probably via gut microbiota [J]. Frontiers in Microbiology, 2020, 11: 1604. doi: 10.3389/fmicb.2020.01604 [74] LIU Y H, LI Y H, XIA Y H, et al. The dysbiosis of gut microbiota caused by low-dose cadmium aggravate the injury of mice liver through increasing intestinal permeability [J]. Microorganisms, 2020, 8(2): 211. doi: 10.3390/microorganisms8020211 [75] NINKOV M, POPOV ALEKSANDROV A, DEMENESKU J, et al. Toxicity of oral cadmium intake: Impact on gut immunity [J]. Toxicology Letters, 2015, 237(2): 89-99. doi: 10.1016/j.toxlet.2015.06.002 [76] TINKOV A A, GRITSENKO V A, SKALNAYA M G, et al. Gut as a target for cadmium toxicity [J]. Environmental Pollution (Barking, Essex:1987), 2018, 235: 429-434. doi: 10.1016/j.envpol.2017.12.114 [77] GAO B, CHI L, MAHBUB R, et al. Multi-omics reveals that lead exposure disturbs gut microbiome development, key metabolites, and metabolic pathways [J]. Chemical Research in Toxicology, 2017, 30(4): 996-1005. doi: 10.1021/acs.chemrestox.6b00401 [78] XIA J Z, JIN C Y, PAN Z H, et al. Chronic exposure to low concentrations of lead induces metabolic disorder and dysbiosis of the gut microbiota in mice [J]. Science of the Total Environment, 2018, 631/632: 439-448. doi: 10.1016/j.scitotenv.2018.03.053 [79] XIA J Z, LU L, JIN C Y, et al. Effects of short term lead exposure on gut microbiota and hepatic metabolism in adult zebrafish [J]. Comparative Biochemistry and Physiology Part C:Toxicology & Pharmacology, 2018, 209: 1-8. [80] ZHAI Q X, LIU Y, WANG C, et al. Lactobacillus plantarum CCFM8661 modulates bile acid enterohepatic circulation and increases lead excretion in mice [J]. Food & Function, 2019, 10(3): 1455-1464. [81] ZHAI Q X, QU D W, FENG S S, et al. Oral supplementation of lead-intolerant intestinal microbes protects against lead (Pb) toxicity in mice [J]. Frontiers in Microbiology, 2020, 10: 3161. doi: 10.3389/fmicb.2019.03161 [82] ZHAI Q X, WANG J, CEN S, et al. Modulation of the gut microbiota by a galactooligosaccharide protects against heavy metal lead accumulation in mice [J]. Food & Function, 2019, 10(6): 3768-3781. [83] XING S C, HUANG C B, MI J D, et al. Bacillus coagulans R11 maintained intestinal villus health and decreased intestinal injury in lead-exposed mice by regulating the intestinal microbiota and influenced the function of faecal microRNAs [J]. Environmental Pollution, 2019, 255: 113139. doi: 10.1016/j.envpol.2019.113139 [84] ZHAO Y P, QIAO R X, ZHANG S Y, et al. Metabolomic profiling reveals the intestinal toxicity of different length of microplastic fibers on zebrafish (Danio rerio) [J]. Journal of Hazardous Materials, 2021, 403: 123663. doi: 10.1016/j.jhazmat.2020.123663 [85] PAITHANKAR J G, SAINI S, DWIVEDI S, et al. Heavy metal associated health hazards: An interplay of oxidative stress and signal transduction [J]. Chemosphere, 2021, 262: 128350. doi: 10.1016/j.chemosphere.2020.128350 [86] AMUNO S, AL KAISSI A, JAMWAL A, et al. Chronic arsenicosis and cadmium exposure in wild snowshoe hares (Lepus americanus) breeding near Yellowknife, Northwest Territories (Canada), part 1: Evaluation of oxidative stress, antioxidant activities and hepatic damage [J]. Science of the Total Environment, 2018, 618: 916-926. doi: 10.1016/j.scitotenv.2017.08.278 [87] DONG X X, SHULZHENKO N, LEMAITRE J, et al. Arsenic exposure and intestinal microbiota in children from Sirajdikhan, Bangladesh [J]. PLoS One, 2017, 12(12): e0188487. doi: 10.1371/journal.pone.0188487 [88] YarMOHAMMADI A A, BIDGOLI S A, ZIARATI P. Increased urinary arsenic concentration in newly diagnosed type 2 diabetes mellitus: A gender-independent, smoking-dependent exposure biomarker in older adults in Tehran [J]. Environmental Science and Pollution Research, 2021, 28(22): 27769-27777. doi: 10.1007/s11356-020-10261-w [89] DHEER R, PATTERSON J, DUDASH M, et al. Arsenic induces structural and compositional colonic microbiome change and promotes host nitrogen and amino acid metabolism [J]. Toxicology and Applied Pharmacology, 2015, 289(3): 397-408. doi: 10.1016/j.taap.2015.10.020 [90] LU K, ABO R P, SCHLIEPER K A, et al. Arsenic exposure perturbs the gut microbiome and its metabolic profile in mice: An integrated metagenomics and metabolomics analysis [J]. Environmental Health Perspectives, 2014, 122(3): 284-291. doi: 10.1289/ehp.1307429 [91] BRABEC J L, WRIGHT J, LY T, et al. Arsenic disturbs the gut microbiome of individuals in a disadvantaged community in Nepal [J]. Heliyon, 2020, 6(1): e03313. doi: 10.1016/j.heliyon.2020.e03313 [92] HOEN A G, MADAN J C, LI Z G, et al. Sex-specific associations of infants’gut microbiome with arsenic exposure in a US population [J]. Scientific Reports, 2018, 8: 12627. doi: 10.1038/s41598-018-30581-9 [93] CORYELL M, McALPINE M, PINKHAM N V, et al. The gut microbiome is required for full protection against acute arsenic toxicity in mouse models [J]. Nature Communications, 2018, 9: 5424. doi: 10.1038/s41467-018-07803-9 [94] ZHAO Y L, ZHOU C M, WU C, et al. Subchronic oral mercury caused intestinal injury and changed gut microbiota in mice [J]. Science of the Total Environment, 2020, 721: 137639. doi: 10.1016/j.scitotenv.2020.137639 [95] 张进. 铜和汞对小鼠盲肠、直肠微生物多样性的影响 [D]. 南昌: 江西农业大学, 2017. ZHANG J. Effects of copper and mercury on microbiota diversity in cecum and rectum of mice [D]. Nanchang: Jiangxi Agricultural University, 2017 (in Chinese).
[96] LIN X Y, ZHAO J T, ZHANG W, et al. Acute oral methylmercury exposure perturbs the gut microbiome and alters gut-brain axis related metabolites in rats [J]. Ecotoxicology and Environmental Safety, 2020, 190: 110130. doi: 10.1016/j.ecoenv.2019.110130 [97] NIELSEN K M, ZHANG Y, CURRAN T E, et al. Alterations to the intestinal microbiome and metabolome of Pimephales promelas and mus musculus following exposure to dietary methylmercury [J]. Environmental Science & Technology, 2018, 52(15): 8774-8784. [98] ROTHENBERG S E, WAGNER C L, HAMIDI B, et al. Longitudinal changes during pregnancy in gut microbiota and methylmercury biomarkers, and reversal of microbe-exposure correlations [J]. Environmental Research, 2019, 172: 700-712. doi: 10.1016/j.envres.2019.01.014 [99] SEKI N, AKIYAMA M, YAMAKAWA H, et al. Adverse effects of methylmercury on gut bacteria and accelerated accumulation of mercury in organs due to disruption of gut microbiota [J]. The Journal of Toxicological Sciences, 2021, 46(2): 91-97. doi: 10.2131/jts.46.91 [100] CHANG J, ZHOU Y, WANG Q, et al. Plant components can reduce methylmercury toxication: A mini-review [J]. Biochimica et Biophysica Acta (BBA) - General Subjects, 2019, 1863(12): 129290. doi: 10.1016/j.bbagen.2019.01.012 [101] JIANG X P, GU S S, LIU D, et al. Lactobacillus brevis 23017 relieves mercury toxicity in the colon by modulation of oxidative stress and inflammation through the interplay of MAPK and NF-κB signaling cascades [J]. Frontiers in Microbiology, 2018, 9: 2425. doi: 10.3389/fmicb.2018.02425 [102] RICHARDSON J B, DANCY B C R, HORTON C L, et al. Exposure to toxic metals triggers unique responses from the rat gut microbiota [J]. Scientific Reports, 2018, 8: 6578. doi: 10.1038/s41598-018-24931-w [103] ZHANG Z C, CAO H Y, SONG N, et al. Long-term hexavalent chromium exposure facilitates colorectal cancer in mice associated with changes in gut microbiota composition [J]. Food and Chemical Toxicology, 2020, 138: 111237. doi: 10.1016/j.fct.2020.111237 [104] CHI L, GAO B, Bian X M, et al. Manganese-induced sex-specific gut microbiome perturbations in C57BL/6 mice [J]. Toxicology and Applied Pharmacology, 2017, 331: 142-153. doi: 10.1016/j.taap.2017.06.008 [105] MENG X L, LI S, QIN C B, et al. Intestinal microbiota and lipid metabolism responses in the common carp (Cyprinus carpio L. ) following copper exposure [J]. Ecotoxicology and Environmental Safety, 2018, 160: 257-264. doi: 10.1016/j.ecoenv.2018.05.050 [106] ZHANG Y, ZHANG P J, SHANG X C, et al. Exposure of lead on intestinal structural integrity and the diversity of gut microbiota of common carp [J]. Comparative Biochemistry and Physiology Part C:Toxicology & Pharmacology, 2021, 239: 108877. [107] QIU Y L, CHEN X S, YAN X Y et al. Gut microbiota perturbations and neurodevelopmental impacts in offspring rats concurrently exposure to inorganic arsenic and fluoride [J]. Environment International, 2020, 140: 105763. doi: 10.1016/j.envint.2020.105763 [108] DAHAN D, JUDE B A, LAMENDELLA R, et al. Exposure to arsenic alters the microbiome of larval zebrafish [J]. Frontiers in Microbiology, 2018, 9: 1323. doi: 10.3389/fmicb.2018.01323 [109] WANG H T, DING J, XIONG C, et al. Exposure to microplastics lowers arsenic accumulation and alters gut bacterial communities of earthworm Metaphire californica [J]. Environmental Pollution, 2019, 251: 110-116. doi: 10.1016/j.envpol.2019.04.054 [110] ZHAO Y L, ZHOU C M, GUO X Q, et al. Exposed to mercury-induced oxidative stress, changes of intestinal microflora, and association between them in mice [J]. Biological Trace Element Research, 2021, 199(5): 1900-1907. doi: 10.1007/s12011-020-02300-x [111] ZHOU C M, XU P Z, HUANG C, et al. Effects of subchronic exposure of mercuric chloride on intestinal histology and microbiota in the cecum of chicken [J]. Ecotoxicology and Environmental Safety, 2020, 188: 109920. doi: 10.1016/j.ecoenv.2019.109920 [112] ZHU J, TANG L, QIAO S L, et al. Low-dose methylmercury exposure impairs the locomotor activity of zebrafish: Role of intestinal inositol metabolism [J]. Environmental Research, 2020, 190: 110020. doi: 10.1016/j.envres.2020.110020 [113] RICHARDSON S D, TERNES T A. Water analysis: Emerging contaminants and current issues [J]. Analytical Chemistry, 2005, 77(12): 3807-3838. doi: 10.1021/ac058022x [114] MUIR D C G, HOWARD P H. Are there other persistent organic pollutants? A challenge for environmental chemists [J]. Environmental Science & Technology, 2006, 40(23): 7157-7166. [115] 王燕飞, 蒋京呈, 胡俊杰, 等. 新污染物治理国际经验与启示[J]. 环境保护, 2022, 50(20): 61-66. WANG Y F, JIANG J C, HU J J, et al. International experience and enlightenment of controlling emerging contaminants, Environmental Protection, 2022, 50(20): 61-66 (in Chinese).
[116] WANG Z Y, DeWITT J C, HIGGINS C P, et al. A never-ending story of per- and polyfluoroalkyl substances (PFASs)? [J]. Environmental Science & Technology, 2017, 51(5): 2508-2518. [117] LIU X D, LI L Q, GU L, et al. Distribution and release of perfluorinated compounds (PFCs) in water-sediment systems: The effect of confluence channels [J]. Science of the Total Environment, 2021, 775: 145720. doi: 10.1016/j.scitotenv.2021.145720 [118] CONDER J M, HOKE R A, de WOLF W, et al. Are PFCAs bioaccumulative? A critical review and comparison with regulatory criteria and persistent lipophilic compounds [J]. Environmental Science & Technology, 2008, 42(4): 995-1003. [119] RASHID F, AHMAD S, IRUDAYARAJ J M K. Effect of perfluorooctanoic acid on the epigenetic and tight junction genes of the mouse intestine [J]. Toxics, 2020, 8(3): 64. doi: 10.3390/toxics8030064 [120] WANG G, PAN R L, LIANG X, et al. Perfluorooctanoic acid-induced liver injury is potentially associated with gut microbiota dysbiosis [J]. Chemosphere, 2021, 266: 129004. doi: 10.1016/j.chemosphere.2020.129004 [121] MENG X, LI S, LI Y, et al. Gut microbiota’s relationship with liver disease and role in hepatoprotection by dietary natural products and probiotics [J]. Nutrients, 2018, 10(10): 1457. doi: 10.3390/nu10101457 [122] SHI L C, ZHENG J J, YAN S K, et al. Exposure to perfluorooctanoic acid induces cognitive deficits via altering gut microbiota composition, impairing intestinal barrier integrity, and causing inflammation in gut and brain [J]. Journal of Agricultural and Food Chemistry, 2020, 68(47): 13916-13928. doi: 10.1021/acs.jafc.0c05834 [123] XU C, JIANG Z Y, LIU Q, et al. Estrogen receptor beta mediates hepatotoxicity induced by perfluorooctane sulfonate in mouse [J]. Environmental Science and Pollution Research, 2017, 24(15): 13414-13423. doi: 10.1007/s11356-017-8943-3 [124] LAI K P, NG A H M, WAN H T, et al. Dietary exposure to the environmental chemical, PFOS on the diversity of gut microbiota, associated with the development of metabolic syndrome [J]. Frontiers in Microbiology, 2018, 9: 2552. doi: 10.3389/fmicb.2018.02552 [125] WANG L J, TANG L, FENG Y M, et al. A purified membrane protein from Akkermansia muciniphila or the pasteurised bacterium blunts colitis associated tumourigenesis by modulation of CD8+ T cells in mice [J]. Gut, 2020, 69(11): 1988-1997. doi: 10.1136/gutjnl-2019-320105 [126] ZHANG L M, RIMAL B, NICHOLS R G, et al. Perfluorooctane sulfonate alters gut microbiota-host metabolic homeostasis in mice [J]. Toxicology, 2020, 431: 152365. doi: 10.1016/j.tox.2020.152365 [127] BAO Y X, QU Y X, HUANG J, et al. First assessment on degradability of sodium p-perfluorous nonenoxybenzene sulfonate (OBS), a high volume alternative to perfluorooctane sulfonate in fire-fighting foams and oil production agents in China [J]. RSC Advances, 2017, 7(74): 46948-46957. doi: 10.1039/C7RA09728J [128] WANG W, MI X, SHI H L, et al. Adsorption behaviour and mechanism of the PFOS substitute OBS (sodium p-perfluorous nonenoxybenzene sulfonate) on activated carbon [J]. Royal Society Open Science, 2019, 6(9): 191069. doi: 10.1098/rsos.191069 [129] WANG C Y, ZHAO Y, JIN Y X. The emerging PFOS alternative OBS exposure induced gut microbiota dysbiosis and hepatic metabolism disorder in adult zebrafish [J]. Comparative Biochemistry and Physiology Part C:Toxicology & Pharmacology, 2020, 230: 108703. [130] CHEN L G, LAM J C W, HU C Y, et al. Perfluorobutanesulfonate exposure causes durable and transgenerational dysbiosis of gut microbiota in marine medaka [J]. Environmental Science & Technology Letters, 2018, 5(12): 731-738. [131] ZHOU J F, SHU R N, YU C N, et al. Exposure to low concentration of trifluoromethanesulfonic acid induces the disorders of liver lipid metabolism and gut microbiota in mice [J]. Chemosphere, 2020, 258: 127255. doi: 10.1016/j.chemosphere.2020.127255 [132] ELBERT A, HAAS M, SPRINGER B, et al. Applied aspects of neonicotinoid uses in crop protection [J]. Pest Management Science, 2008, 64(11): 1099-1105. doi: 10.1002/ps.1616 [133] 张敏恒, 赵平, 严秋旭, 等. 新烟碱类杀虫剂市场与环境影响 [J]. 农药, 2012, 51(12): 859-862,900. doi: 10.16820/j.cnki.1006-0413.2012.12.001 ZHANG M H, ZHAO P, YAN Q X, et al. The market and environmental impact of the neonicotinoid insecticides [J]. Agrochemicals, 2012, 51(12): 859-862,900(in Chinese). doi: 10.16820/j.cnki.1006-0413.2012.12.001
[134] 张国生, 侯广新. 烟碱类杀虫剂的应用、开发现状及展望 [J]. 农药科学与管理, 2004, 25(3): 22-26. doi: 10.3969/j.issn.1002-5480.2004.03.009 ZHANG G S, HOU G X. Present status of development and prospect of nicotinoid insecticides [J]. Pesticide Science and Administration, 2004, 25(3): 22-26(in Chinese). doi: 10.3969/j.issn.1002-5480.2004.03.009
[135] WINTERMANTEL D, LOCKE B, ANDERSSON G K S, et al. Field-level clothianidin exposure affects bumblebees but generally not their pathogens [J]. Nature Communications, 2018, 9: 5446. doi: 10.1038/s41467-018-07914-3 [136] BLACQUIÈRE T, SMAGGHE G, van GESTEL C A M, et al. Neonicotinoids in bees: A review on concentrations, side-effects and risk assessment [J]. Ecotoxicology, 2012, 21(4): 973-992. doi: 10.1007/s10646-012-0863-x [137] HALLMANN C A, FOPPEN R P B, van TURNHOUT C A M, et al. Declines in insectivorous birds are associated with high neonicotinoid concentrations [J]. Nature, 2014, 511(7509): 341-343. doi: 10.1038/nature13531 [138] LI Y J, MIAO R Q, KHANNA M. Neonicotinoids and decline in bird biodiversity in the United States [J]. Nature Sustainability, 2020, 3(12): 1027-1035. doi: 10.1038/s41893-020-0582-x [139] KIM J H, KIM D, MOON S M, et al. Associations of lifestyle factors with phthalate metabolites, bisphenol A, parabens, and triclosan concentrations in breast milk of Korean mothers [J]. Chemosphere, 2020, 249: 126149. doi: 10.1016/j.chemosphere.2020.126149 [140] HAO W J, ZHU H Y, CHEN J N, et al. Wild melon seed oil reduces plasma cholesterol and modulates gut microbiota in hypercholesterolemic hamsters [J]. Journal of Agricultural and Food Chemistry, 2020, 68(7): 2071-2081. doi: 10.1021/acs.jafc.9b07302 [141] YAN S, MENG Z Y, TIAN S N, et al. Neonicotinoid insecticides exposure cause amino acid metabolism disorders, lipid accumulation and oxidative stress in ICR mice [J]. Chemosphere, 2020, 246: 125661. doi: 10.1016/j.chemosphere.2019.125661 [142] YANG G L, YUAN X L, JIN C Y, et al. Imidacloprid disturbed the gut barrier function and interfered with bile acids metabolism in mice [J]. Environmental Pollution (Barking, Essex: 1987), 2020, 266(Pt 1): 115290 [143] IKEGAMI T, HONDA A. Reciprocal interactions between bile acids and gut microbiota in human liver diseases [J]. Hepatology Research, 2018, 48(1): 15-27. doi: 10.1111/hepr.13001 [144] ONARU K, OHNO S, KUBO S, et al. Immunotoxicity evaluation by subchronic oral administration of clothianidin in Sprague-Dawley rats [J]. The Journal of Veterinary Medical Science, 2020, 82(3): 360-372. doi: 10.1292/jvms.19-0689 [145] GILL R J, RAMOS-RODRIGUEZ O, RAINE N E. Combined pesticide exposure severely affects individual- and colony-level traits in bees [J]. Nature, 2012, 491(7422): 105-108. doi: 10.1038/nature11585 [146] HIRANO T, YANAI S, OMOTEHARA T, et al. The combined effect of clothianidin and environmental stress on the behavioral and reproductive function in male mice [J]. The Journal of Veterinary Medical Science, 2015, 77(10): 1207-1215. doi: 10.1292/jvms.15-0188 [147] FROMME H, KÜCHLER T, OTTO T, et al. Occurrence of phthalates and bisphenol A and F in the environment [J]. Water Research, 2002, 36(6): 1429-1438. doi: 10.1016/S0043-1354(01)00367-0 [148] ROCHESTER J R. Bisphenol A and human health: A review of the literature [J]. Reproductive Toxicology , 2013, 42: 132-155. doi: 10.1016/j.reprotox.2013.08.008 [149] ACCONCIA F, PALLOTTINI V, MARINO M. Molecular mechanisms of action of BPA [J]. Dose-Response, 2015, 13(4): 1559325815610582 [150] FENG D, ZHANG H M, JIANG X, et al. Bisphenol A exposure induces gut microbiota dysbiosis and consequent activation of gut-liver axis leading to hepatic steatosis in CD-1 mice [J]. Environmental Pollution (Barking, Essex: 1987), 2020, 265(Pt A): 114880. [151] DeLUCA J A, ALLRED K F, MENON R, et al. Bisphenol-a alters microbiota metabolites derived from aromatic amino acids and worsens disease activity during colitis [J]. Experimental Biology and Medicine, 2018, 243(10): 864-875. doi: 10.1177/1535370218782139 [152] DIAMANTE G, CELY I, ZAMORA Z, et al. Systems toxicogenomics of prenatal low-dose BPA exposure on liver metabolic pathways, gut microbiota, and metabolic health in mice [J]. Environment International, 2021, 146: 106260. doi: 10.1016/j.envint.2020.106260 [153] GASCON M, CASAS M, MORALES E, et al. Prenatal exposure to bisphenol A and phthalates and childhood respiratory tract infections and allergy [J]. The Journal of Allergy and Clinical Immunology, 2015, 135(2): 370-378. doi: 10.1016/j.jaci.2014.09.030 [154] MALAISÉ Y, MÉNARD S, CARTIER C, et al. Consequences of bisphenol a perinatal exposure on immune responses and gut barrier function in mice [J]. Archives of Toxicology, 2018, 92(1): 347-358. doi: 10.1007/s00204-017-2038-2 [155] JAVUREK A B, SPOLLEN W G, JOHNSON S A, et al. Effects of exposure to bisphenol A and ethinyl estradiol on the gut microbiota of parents and their offspring in a rodent model [J]. Gut Microbes, 2016, 7(6): 471-485. doi: 10.1080/19490976.2016.1234657 [156] OISHI K, SATO T, YOKOI W, et al. Effect of probiotics, Bifidobacterium breve and Lactobacillus casei, on bisphenol A exposure in rats [J]. Bioscience, Biotechnology, and Biochemistry, 2008, 72(6): 1409-1415. doi: 10.1271/bbb.70672 [157] 邓媛媛, 王军, 姚佳希, 等. 双酚S暴露对小鼠脑-肠-菌轴神经递质代谢稳态的影响 [J]. 南京医科大学学报(自然科学版), 2020, 40(5): 663-668. DENG Y Y, WANG J, YAO J X, et al. Bisphenol S exposure affected the mice neurotransmitter metabolism homeostasis through the brain-gut-microbiota axis [J]. Journal of Nanjing Medical University (Natural Sciences), 2020, 40(5): 663-668(in Chinese).
[158] WANG W W, RU S G, WANG L L, et al. Bisphenol S induces ectopic angiogenesis in embryos via VEGFR2 signaling, leading to lipid deposition in blood vessels of larval zebrafish [J]. Environmental Science & Technology, 2020, 54(11): 6822-6831. [159] CATRON T R, KEELY S P, BRINKMAN N E, et al. Host developmental toxicity of BPA and BPA alternatives is inversely related to microbiota disruption in zebrafish [J]. Toxicological Sciences, 2019, 167(2): 468-483. doi: 10.1093/toxsci/kfy261 [160] SINGER H, MÜLLER S, TIXIER C, et al. Triclosan: Occurrence and fate of a widely used biocide in the aquatic environment: Field measurements in wastewater treatment plants, surface waters, and lake sediments [J]. Environmental Science & Technology, 2002, 36(23): 4998-5004. [161] ALLMYR M, ADOLFSSON-ERICI M, McLACHLAN M S, et al. Triclosan in plasma and milk from Swedish nursing mothers and their exposure via personal care products [J]. The Science of the Total Environment, 2006, 372(1): 87-93. doi: 10.1016/j.scitotenv.2006.08.007 [162] BAI X Y, ZHANG B, HE Y, et al. Triclosan and triclocarbon in maternal-fetal serum, urine, and amniotic fluid samples and their implication for prenatal exposure [J]. Environmental Pollution, 2020, 266(Pt 1): 115117. [163] CANDACE S B, AMY A R, MALIN N, et al. Effects of triclosan in breast milk on the infant fecal microbiome [J]. Chemosphere, 2018, 203: 467-473. doi: 10.1016/j.chemosphere.2018.03.186 [164] KIKUCHI K, SAIGUSA D, KANEMITSU Y, et al. Gut microbiome-derived phenyl sulfate contributes to albuminuria in diabetic kidney disease [J]. Nature Communications, 2019, 10: 1835. doi: 10.1038/s41467-019-09735-4 [165] MAHALAK K K, FIRRMAN J, LEE J J, et al. Triclosan has a robust, yet reversible impact on human gut microbial composition in vitro [J]. PLoS One, 2020, 15(6): e0234046. doi: 10.1371/journal.pone.0234046 [166] MA Y, GUO Y S, YE H L, et al. Perinatal Triclosan exposure in the rat induces long-term disturbances in metabolism and gut microbiota in adulthood and old age [J]. Environmental Research, 2020, 182: 109004. doi: 10.1016/j.envres.2019.109004 [167] ZHANG H N, LIANG Y S, WU P F, et al. Continuous dermal exposure to triclocarban perturbs the homeostasis of liver-gut axis in mice: Insights from metabolic interactions and microbiome shifts [J]. Environmental Science & Technology, 2021, 55(8): 5117-5127. [168] YANG H X, SANIDAD K Z, WANG W C, et al. Triclocarban exposure exaggerates colitis and colon tumorigenesis: Roles of gut microbiota involved [J]. Gut Microbes, 2020, 12(1): 1690364. doi: 10.1080/19490976.2019.1690364 [169] EERKES-MEDRANO D, THOMPSON R C, ALDRIDGE D C. Microplastics in freshwater systems: A review of the emerging threats, identification of knowledge gaps and prioritisation of research needs [J]. Water Research, 2015, 75: 63-82. doi: 10.1016/j.watres.2015.02.012 [170] DUIS K, COORS A. Microplastics in the aquatic and terrestrial environment: Sources (with a specific focus on personal care products), fate and effects [J]. Environmental Sciences Europe, 2016, 28(1): 2. doi: 10.1186/s12302-015-0069-y [171] WRIGHT S L, KELLY F J. Plastic and human health: A micro issue? [J]. Environmental Science & Technology, 2017, 51(12): 6634-6647. [172] JIN Y X, XIA J Z, PAN Z H, et al. Polystyrene microplastics induce microbiota dysbiosis and inflammation in the gut of adult zebrafish [J]. Environmental Pollution, 2018, 235: 322-329. doi: 10.1016/j.envpol.2017.12.088 [173] FACKELMANN G, SOMMER S. Microplastics and the gut microbiome: How chronically exposed species may suffer from gut dysbiosis [J]. Marine Pollution Bulletin, 2019, 143: 193-203. doi: 10.1016/j.marpolbul.2019.04.030 [174] LEI L L, WU S Y, LU S B, et al. Microplastic particles cause intestinal damage and other adverse effects in zebrafish Danio rerio and nematode Caenorhabditis elegans [J]. The Science of the Total Environment, 2018, 619/620: 1-8. doi: 10.1016/j.scitotenv.2017.11.103 [175] QIAO R X, DENG Y F, ZHANG S H, et al. Accumulation of different shapes of microplastics initiates intestinal injury and gut microbiota dysbiosis in the gut of zebrafish [J]. Chemosphere, 2019, 236: 124334. doi: 10.1016/j.chemosphere.2019.07.065 [176] LU L, WAN Z Q, LUO T, et al. Polystyrene microplastics induce gut microbiota dysbiosis and hepatic lipid metabolism disorder in mice [J]. The Science of the Total Environment, 2018, 631/632: 449-458. doi: 10.1016/j.scitotenv.2018.03.051 [177] JIN Y X, LU L, TU W Q, et al. Impacts of polystyrene microplastic on the gut barrier, microbiota and metabolism of mice [J]. The Science of the Total Environment, 2019, 649: 308-317. doi: 10.1016/j.scitotenv.2018.08.353 [178] DENG Y F, YAN Z H, SHEN R Q, et al. Microplastics release phthalate esters and cause aggravated adverse effects in the mouse gut [J]. Environment International, 2020, 143: 105916. doi: 10.1016/j.envint.2020.105916 [179] SHENG C, ZHANG S H, ZHANG Y. The influence of different polymer types of microplastics on adsorption, accumulation, and toxicity of triclosan in zebrafish [J]. Journal of Hazardous Materials, 2021, 402: 123733. doi: 10.1016/j.jhazmat.2020.123733 [180] SABRI N A, SCHMITT H, van der ZAAN B, et al. Prevalence of antibiotics and antibiotic resistance genes in a wastewater effluent-receiving river in the Netherlands [J]. Journal of Environmental Chemical Engineering, 2020, 8(1): 102245. doi: 10.1016/j.jece.2018.03.004 [181] ANJALI R, SHANTHAKUMAR S. Insights on the current status of occurrence and removal of antibiotics in wastewater by advanced oxidation processes [J]. Journal of Environmental Management, 2019, 246: 51-62. [182] QIAN M R, WU H Z, WANG J M, et al. Occurrence of trace elements and antibiotics in manure-based fertilizers from the Zhejiang Province of China [J]. The Science of the Total Environment, 2016, 559: 174-181. doi: 10.1016/j.scitotenv.2016.03.123 [183] ZHAN J, LIANG Y R, LIU D H, et al. Antibiotics may increase triazine herbicide exposure risk via disturbing gut microbiota [J]. Microbiome, 2018, 6(1): 224. doi: 10.1186/s40168-018-0602-5 [184] HAGAN T, CORTESE M, ROUPHAEL N, et al. Antibiotics-driven gut microbiome perturbation alters immunity to vaccines in humans [J]. Cell, 2019, 178(6): 1313-1328.e13. doi: 10.1016/j.cell.2019.08.010 [185] JERNBERG C, LÖFMARK S, EDLUND C, et al. Long-term ecological impacts of antibiotic administration on the human intestinal microbiota [J]. The ISME Journal, 2007, 1(1): 56-66. doi: 10.1038/ismej.2007.3 [186] BUFFIE C G, JARCHUM I, EQUINDA M, et al. Profound alterations of intestinal microbiota following a single dose of clindamycin results in sustained susceptibility to Clostridium difficile-induced colitis [J]. Infection and Immunity, 2012, 80(1): 62-73. doi: 10.1128/IAI.05496-11 [187] BUFFIE C G, BUCCI V, STEIN R R, et al. Precision microbiome reconstitution restores bile acid mediated resistance to Clostridium difficile [J]. Nature, 2015, 517(7533): 205-208. doi: 10.1038/nature13828 [188] WU Y Q, ZHENG Y F, CHEN Y N, et al. Honey bee (Apis mellifera) gut microbiota promotes host endogenous detoxification capability via regulation of P450 gene expression in the digestive tract [J]. Microbial Biotechnology, 2020, 13(4): 1201-1212. doi: 10.1111/1751-7915.13579 [189] MU Q H, ZHANG H S, LIAO X F, et al. Control of lupus nephritis by changes of gut microbiota [J]. Microbiome, 2017, 5(1): 73. doi: 10.1186/s40168-017-0300-8 [190] GÓMEZ-HURTADO I, MORATALLA A, MOYA-PÉREZ Á, et al. Role of interleukin 10 in norfloxacin prevention of luminal free endotoxin translocation in mice with cirrhosis [J]. Journal of Hepatology, 2014, 61(4): 799-808. doi: 10.1016/j.jhep.2014.05.031 -
 点击查看大图
点击查看大图
计量
- 文章访问数: 4825
- HTML全文浏览数: 4825
- PDF下载数: 123
- 施引文献: 0
