

-
邻苯二甲酸酯类化合物(phthalate esters, PAEs)是邻苯二甲酸酐与醇反应生成的化合物,它是提高聚氯乙烯(polyvinyl chloride,PVC)弹性的重要添加剂,其中邻苯二甲酸二(2-乙基己基)酯(Di(2-ethylhexyl)phthalate,DEHP)是目前使用量最大的一种邻苯二甲酸酯,用量高达80%[1]. DEHP广泛应用于儿童玩具、塑料包装、化妆品及各类医疗器械[2-3]. DEHP的广泛使用导致了不可避免的环境释放以及人体摄入. 大量实验指出我国人群DEHP的暴露剂量约为11—116 μg·kg−1·d−1,接近DEHP的每日可耐受摄入量(TDI)(20—140 μg·kg−1·d−1),表明 DEHP对人群的健康构成重大危害[4].
DEHP具有内分泌干扰效应,可以诱发生殖发育毒性、肝脏毒性、胚胎毒性等多种毒性[5-6]. DEHP通过消化道进入人体后在胃肠道的脂肪酶作用下水解成初级代谢物乙基己醇(2-EH)和邻苯二甲酸单乙基己基酯(mono-ethylhexyl phthalate,MEHP),再通过尿液排出体外[7]. 通过对尿液的单酯类物检测发现,原尿和酶解后的尿液中 MEHP 的检出率最高,平均检出率超过60% [8]. 此外,试验结果表明DEHP的毒性主要来源于其代谢物 MEHP,其毒性作用高达DEHP的10倍[9]. 同时,MEHP的动物实验表明,其主要分布在肾脏、膀胱和肝脏,其中肝脏为MEHP最主要的靶器官[10]. Thomas等的研究中,大鼠口服500 mg·kg−1 DEHP,30 min后,肝脏MEHP水平为12.5 mg·g−1 [11]. MEHP毒性作用主要体现在增加肝脏亲脂活性,导致肝脏中出现脂肪堆积,继而引发肝细胞脂肪变性[12].
肝脏在体内发挥着外源物质解毒和代谢脂类物质的主要作用. 肝脏内游离脂肪酸增加和甘油三酯沉积是肝脏脂质代谢紊乱的主要表现. 近年来,全球肥胖病与非酒精性脂肪肝患病率快速上升. 体内体外实验提示脂肪代谢紊乱会增加肥胖、糖尿病、非酒精性脂肪肝病的患病危险度. 流行病学和毒理学研究报告称,接触DEHP会影响机体脂肪代谢,从而促进肥胖[13]. 体内实验表明大鼠用DEHP处理后,肝脏重量增加,主要机制可能与肝代谢酶改变有关[14]. 有体外实验表明,MEHP处理HepG2细胞后,激活了PPARα使脂肪酸的氧化分解受到抑制[9],导致肝细胞内的脂质堆积并造成肝脏损伤. 在本课题组的前期研究发现,MEHP 处理后的HepG2 细胞的乙酰辅酶 A 羧化酶(acetyl-CoA carboxylase, ACC)的亚型ACC1蛋白表达水平增加,最终使肝脏细胞中脂肪酸合成增加. 这些结果表明,MEHP 可能通过影响脂质合成相关基因或蛋白的表达,从而导致肝细胞脂肪代谢出现紊乱. 为此,本实验以HepG2细胞为实验对象,通过MEHP染毒,观察细胞内脂质代谢情况,并通过基因芯片高通量筛查差异基因,探讨MEHP对体外脂质合成的影响及其可能的作用机制.
-
从中国科学院上海生命研究院细胞资源中心购买HepG2细胞. 细胞培养基含 10%胎牛血清、1%青霉素庆大霉素双抗. 细胞培养条件为37 ℃、5% CO2. 取指数生长期的细胞进行染毒,参考相关文献[9]及我国人群接触水平设置染毒浓度. 共设置5个染毒浓度 :阴性对照(完全培养基)、阳性对照(1 mmol·L−1油酸)、0.01 、1 、10 μmol·L−1MEHP. 染毒48 h后进行相关实验.
-
为了研究细胞中脂滴的蓄积,用油红O法对HepG2细胞进行染色. 5个实验组的细胞染毒48 h后进行染色. 主要步骤为:弃去孔中废弃培养基,用 PBS 漂洗细胞,4%(质量分数)多聚甲醛固定细胞,油红O染色30 min,60%(体积分数)异丙醇洗去多余染料,苏木精复染细胞核10 s. 用显微镜获取图像并观察细胞内脂滴情况.
-
用1 μmol·L−1 MEHP处理的细胞进行芯片分析,Agilent (人)表达谱芯片由博奥晶典公司完成. 步骤如下:以待检测样品的 total RNA 为起始,进行体外扩增和荧光标记,然后用Oligo(dT)Primer 引物合成cRNA并进行纯化和反转录得到cDNA. 最后用 Klenow Fragment 酶合成带有荧光基团的DNA 进行芯片杂交. 用 Feature Extraction 提数软件对芯片杂交扫描后的图片数据进行处理分析.
-
通过Rstudio limma包将芯片基因的数据进行处理,以表格格式导出并进一步筛选差异表达基因. 差异表达基因(DEG)定义为差异倍数 FC大于 1.5或小于0.67且 P值小于 0.05的基因.
-
将上述分析得出的DEG导入 DAVID在线数据库(
https://david.ncifcrf.gov/tools.jsp ),进行GO分析. 通过对显著表达的基因进行功能注释分析,包括生物学过程(BP)、细胞组成(CC)和分子功能(MF),进而了解差异表达基因的生物学意义. 以 P<0.05认为具有显著性. -
按照1.1节对处于指数生长期的细胞进行染毒,每个实验组设置6个平行对照,染毒48 h后提取RNA. 细胞样品用Trizol试剂裂解,提取总RNA. 用超微量分光光度计测定RNA的含量和纯度. 将符合条件的RNA (A260/A230>2.0,和A260/A280=1.8—2.0)样本用反转录试剂盒将RNA反转录为cDNA备用.
从具有显著性且参与脂质代谢过程的差异基因中选取丙酮酸脱氢酶磷酸酶催化亚基2(PDP2)、磷脂酶A2组 IVE(PLA2G4E)、脂肪酸去饱和酶 6(FADS6)、Q型蛋白酪氨酸磷酸酶受体(PTPRQ)、CD28、甾醇-C5-去饱和酶(SC5D)、甲羟戊酸二磷酸脱羧酶(MVD)作为目的基因,检测mRNA表达水平(荧光定量PCR仪为Line Gene 9600). 引物信息如表1所示. 以GAPDH为内参,数据分析时取Ct值的平均值,用相对定量法2−△△ct法计算目的基因表达的变化情况. 采用 SPSS 26.0 软件对数据进行统计学分析,多组间比较使用单因素方差分析,事后比较用LSD 检验进行,P<0.05认为具有统计学意义.
-
将细胞脂质代谢通路的基因输入 GEPIA 在线分析网站(http://gepia.cancer-pku.cn/),分析其在肝癌组织中的表达. P<0.05认为具有统计学意义.
-
如图1所示,阴性对照组细胞有明显的边界,细胞内没有脂滴存在. 油酸染毒的细胞可见大量明显着色的脂滴,主要分布在细胞膜内侧区域,大小不等. 与阴性对照组相比,0.01 μmol·L−1 MEHP剂量组未见明显改变;1、10 μmol·L−1 MEHP剂量组可观察到少量红色脂滴,且随着染毒浓度的增大,脂滴的数量也逐渐增加,暂时未见对细胞边界的影响. 在前期实验中本课题组对MEHP染毒的HepG2细胞中的甘油三酯(TG)含量进行了定量检测, TG含量结果与油红O染色结果相一致,提示较高剂量 MEHP 暴露可提高细胞中的脂质水平[8]. 这一结果表明, MEHP暴露后,导致HepG2细胞中的甘油三酯蓄积,从而引起肝细胞中的脂质沉积.
-
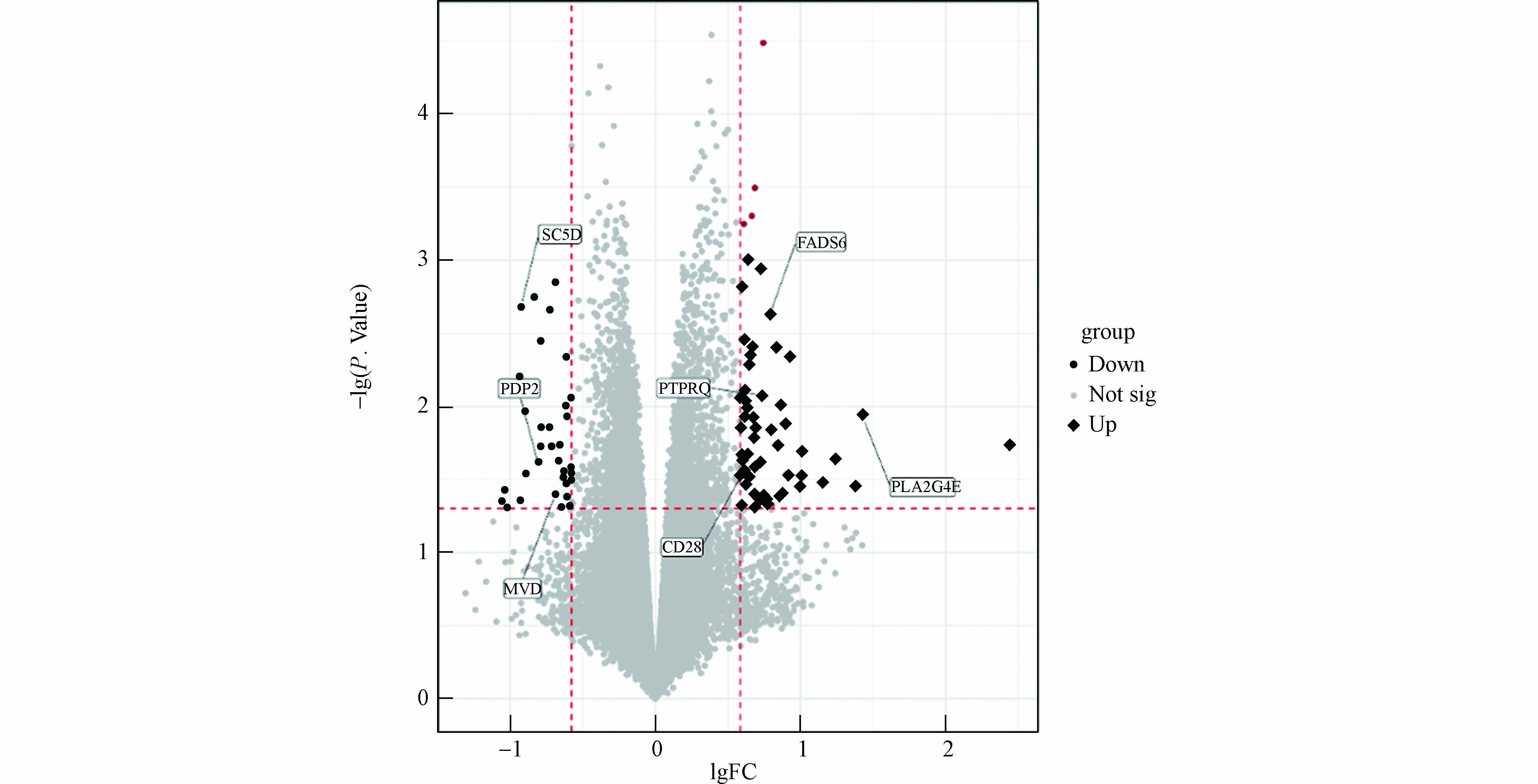
如图2所示,根据P<0.05,FC>1.5或<0.67的筛选标准,共筛选出93个差异表达基因(表2),其中上调的基因57个,下调的基因36个.
-
通过对差异基因进行GO分析探讨差异基因所涉及的生物学过程,包括基因的3个部分:分子功能(MF)、细胞组分(CC)、生物过程(BP). 结果显示,可以识别的基因一共有48个,显著富集的GO条目一共有13条(P<0.05),其中生物过程7条(图3). 前5条通路及具体信息见表3,主要包括细胞脂质代谢过程(cellular lipid metabolic process)、T细胞阴性选择(negative T cell selection)、跨膜运输(transmembrane transport)、跨膜转运的调控(regulation of transmembrane transport)、先天免疫反应(innate immune response);分子功能2条,包括被动跨膜转运活性(passive transmembrane transporter activity)、基质特异性跨膜转运蛋白活性(substrate-specific transmembrane transporter activity);细胞组分4条,包括膜的外部成分(extrinsic component of membrane)、等离子体膜(plasma membrane)、质膜部分(plasma membrane part).GO 分析表明, 最显著激活的通路是细胞脂质代谢通路,他由上调基因 FADS6、PTPRQ、CD28、 PLA2G4E 和下调基因 SC5D、PDP2、MVD 组成.
-
MEHP染毒后相关基因mRNA表达水平的变化如图4所示. 与阴性对照组相比,基因FADS6 在1 μmol·L−1 MEHP染毒样品内基因表达水平明显增加(P<0.05);基因SC5D、PDP2在1 μmol·L−1 MEHP剂量组基因表达水平明显降低(P<0.05),MVD在10 μmol·L−1 MEHP剂量组基因表达水平明显降低(P<0.05). 其中,基因SC5D、PDP2、MVD、FADS6的mRNA表达水平与芯片结果表达一致,基因PTPRQ、CD28、PLA2G4E结果不一致.
本研究通过转录组数据分析探讨MEHP在HepG2细胞脂质代谢过程中发挥作用的基因,发现细胞脂质代谢通路上的基因FADS6、MVD、SC5D、PLA2G4E在代谢过程中起重要作用. 脂肪酸去饱和酶(FADS6)在多不饱和脂肪酸(PUFA)的合成途径中起关键作用,参与了PUFA合成过程中的限速步骤,并调节PUFA的代谢通量[15]. 据报道,FADS6的活性是调节生物体中多不饱和脂肪酸比例的主要因素,其活性改变会影响一些疾病,如炎症和肿瘤发生,2型糖尿病,心血管疾病,代谢紊乱和神经精神疾病[16]. Montell的研究发现,含不饱和脂肪酸的二脂酰甘油(DAG)比饱和脂肪酸的DAG对二酰基甘油酰基转移酶(DGAT)有更强的亲和力,即肝脏合成 TG 首先利用不饱和脂肪酸, 更容易导致甘油三酯积累,而饱和脂肪酸多以 DAG 的形式蓄积[17].
本实验富集分析与PCR的结果显示,FADS6的mRNA表达水平升高,促进了HepG2细胞中多不饱和脂肪酸合成,这可能是HepG2细胞中甘油三酯合成增多进而出现脂肪堆积的原因. 甲羟戊酸二磷酸脱羧酶(MVD)参与胆固醇生物合成过程中的早期步骤. 通过Jump实验中转录组数据的KEGG分析显示,MVD参与SREBP激活基因表达、SREBP调节胆固醇生物合成、脂类和脂蛋白代谢等胆固醇代谢相关的通路,MVD的变化可调控乙酰乙酰-辅酶A的变化,进而改变)胆固醇调节元件结合蛋白(SREBP)的表达水平[18],已有研究证明SREBP的上调导致肝细胞内胆固醇合成增加 [19] . 甾醇-C5-去饱和酶(SC5D)是一种胆固醇生物合成酶,它催化乳甾醇(lathosterol)转化为7-脱氢胆固醇(7-dehydrocholesterol),参与胆固醇合成中的酶促反应通路[20]. 有文献报道SC5D基因缺陷会引起乳甾醇症(Lathosterolosis),一种罕见的常染色体隐性胆固醇生物合成障碍疾病[21]. SC5D基因在HepG2细胞中表达水平的改变导致细胞内胆固醇合成发生紊乱. 磷脂酶A2(PLA2)是一种参与脂蛋白代谢和炎症途径的酶,所产生的脂质介质对细胞代谢起重要调控作用. 有文献报道NAFLD病人血浆中PLA2水平和肝脏的脂肪变性程度呈显著正相关,这说明PLA2可能参与肝细胞代谢过程中[22]. 有研究证实PLA2过表达的肝脏甘油三酯(TG)含量显著增加,同时增加趋势与PLA2的表达趋势相一致[23]. PLA2G4E作为PLA2的一种,对肝脏细胞的代谢也有调控作用. 本研究中发现,芯片结果中PLA2G4E表达水平升高,提示PLA2G4E是MEHP诱导甘油三脂含量增加,扰乱肝细胞脂质代谢的原因之一.
-
利用在线分析工具GEPIA分析以上基因在肝细胞癌组织的表达水平,如图5所示. 这7个基因在肝细胞癌组织中的表达变化对比正常组织没有显著性. 但是以上基因在HepG2肝癌细胞脂质代谢过程中的表达水平均具有明显改变,导致肝癌细胞脂质代谢紊乱、脂肪堆积.
非酒精性脂肪性肝病(nonalcoholic fatty liver disease,NAFLD)是一种可逆性的疾病,但随着研究的不断深入, NAFLD的发病范围由单纯脂肪肝、脂肪肝、肝硬化发展到原发性肝细胞癌[24]. 多项研究证明NAFLD可逐渐发展为非酒精性脂肪性肝炎(Nonalcoholic steatohepatitis,NASH),从而导致肝硬化[25],而肝硬化是除HBV、HCV引起原发性肝癌的另一主要原因. 因此假如高龄、胰岛素抵抗、肥胖等危险因素持续存在,NAFLD最终可引起肝癌. 同时Iris [26]的动物实验和Ertle[27] 的研究证实,NAFLD可不经肝硬化阶段发展为原发性肝癌. 由此可知,虽然以上基因在肝癌组织中的表达没有明显变化,但是对肝癌的前身——NAFLD发生发展具有重要作用.
-
本研究以HepG2细胞为研究对象,结合油红O染色以及基因芯片等技术,发现MEHP暴露影响了细胞脂质代谢等信号通路,改变了关键因子FADS6、MVD、SC5D、PLA2G4E的表达水平,导致肝脏细胞中甘油三酯与胆固醇的合成增加,肝脏细胞出现脂肪蓄积. 综上所述,MEHP暴露可以通过改变细胞代谢相关基因引起肝脏细胞内脂质代谢紊乱.
邻苯二甲酸单乙基己基酯暴露对肝脏脂质代谢的影响及调控机制
Effects of mono- (2-ethylhexyl) phthalate exposure on liver lipid metabolism and its regulatory mechanism
-
摘要: 邻苯二甲酸二(2-乙基己基)酯(Di(2-ethylhexyl) phthalate,DEHP)是一种重要的内分泌干扰物,对神经系统、生殖系统、内分泌系统等都具有特定的损伤作用. DEHP被机体摄入后,在肝脏和肠道中被消化酶代谢为邻苯二甲酸单(2-乙基己基)酯(mono-(2-ethylhexyl)phthalate,MEHP)以发挥毒性作用. 本研究采用体外HepG2暴露结合mRNA芯片以及生信分析的方法,探讨MEHP暴露对HepG2细胞脂质代谢的影响. 首先,采用油红O 染色法观察细胞内脂滴蓄积情况,发现MEHP暴露能够以浓度依赖性的方式增加HepG2细胞内脂质蓄积. 为了探究其可能的机制,采用Agilent(人)表达谱芯片分析并筛选差异基因,共筛选出93个差异表达基因,其中上调基因57个,下调基因36个. 在此基础上,通过DAVID在线数据库对差异表达基因进行基因功能富集分析,发现其主要参与细胞脂质代谢过程(cellular lipid metabolic process)、跨膜运输(transmembrane transport)、跨膜转运的调控(regulation of transmembrane transport)等生物学过程. 随后,通过qRT-PCR法对关键基因的mRNA表达水平进行验证. 最后,通过 GEPIA 在线验证上述基因在肝细胞癌(LIHC)中的表达. 综上,MEHP暴露能够影响细胞脂质代谢通路中的基因FADS6、MVD、SC5D、PLA2G4E在HepG2细胞中的表达,在一定程度上促进了肝脏细胞中的脂肪蓄积,从而使肝细胞内脂质代谢紊乱.
-
关键词:
- 邻苯二甲酸单乙基己基酯 /
- HepG2 细胞 /
- 脂肪蓄积 /
- 生信分析.
Abstract: Di(2-ethylhexyl) phthalate (DEHP) is an important endocrine disruptor, which has specific damage to nervous system, reproductive system and endocrine system. DEHP is metabolized by digestive enzymes into mono-(2-ethylhexyl)phthalate (MEHP) in liver and intestine , so as to produce toxic effects on human health. HepG2 was exposed to MEHP and then mRNA microarray and bioassay was used to investigate the effects of MEHP exposure on lipid metabolism of HepG2. First, oil red O staining was used to observe the accumulation of lipid droplets in HepG2 cells. It was found that MEHP exposure could increase the accumulation of lipid droplets in HepG2 cells. In order to explore the possible mechanism, Agilent (human) expression microarray was used to analyze and screen differentially expressed genes. A total of 93 differentially expressed genes were screened, including 57 up-regulated genes and 36 down-regulated genes. In addition,gene function enrichment analysis of differentially expressed genes was conducted through DAVID online database. It was found that these differentially expressed genes were mainly involved in the regulation of cellular lipid metabolic process, transmembrane transport and transmembrane transport Transport and other biological processes. Then, the mRNA expression levels of key genes were verified by qRT-PCR. Finally, GEPIA was used to verify the expression of these genes in Liver hepatocellular carcinoma(LIHC). In conclusion, MEHP exposure can affect the expression of FADS6、MVD、SC5D and PLA2G4E genes in cellular lipid metabolism pathway in liver, and promote the accumulation of fat in HepG2 cells, thus leading to the disorder of lipid metabolism in liver cells.-
Key words:
- mono- (2-ethylhexyl) phthalate /
- HepG2 cells /
- fat storage /
- raw letter analysis.
-

-

图 1 不同浓度MEHP染毒对HepG2细胞内脂肪积累的影响(×400)
Figure 1. Effects of different concentrations of MEHP on lipid accumulation in HepG2 cells(×400)

图 3 DAVID分析中生物过程通路的GO条目与基因
Figure 3. Chord plot depicting the relationship between genes and GO terms of biological process
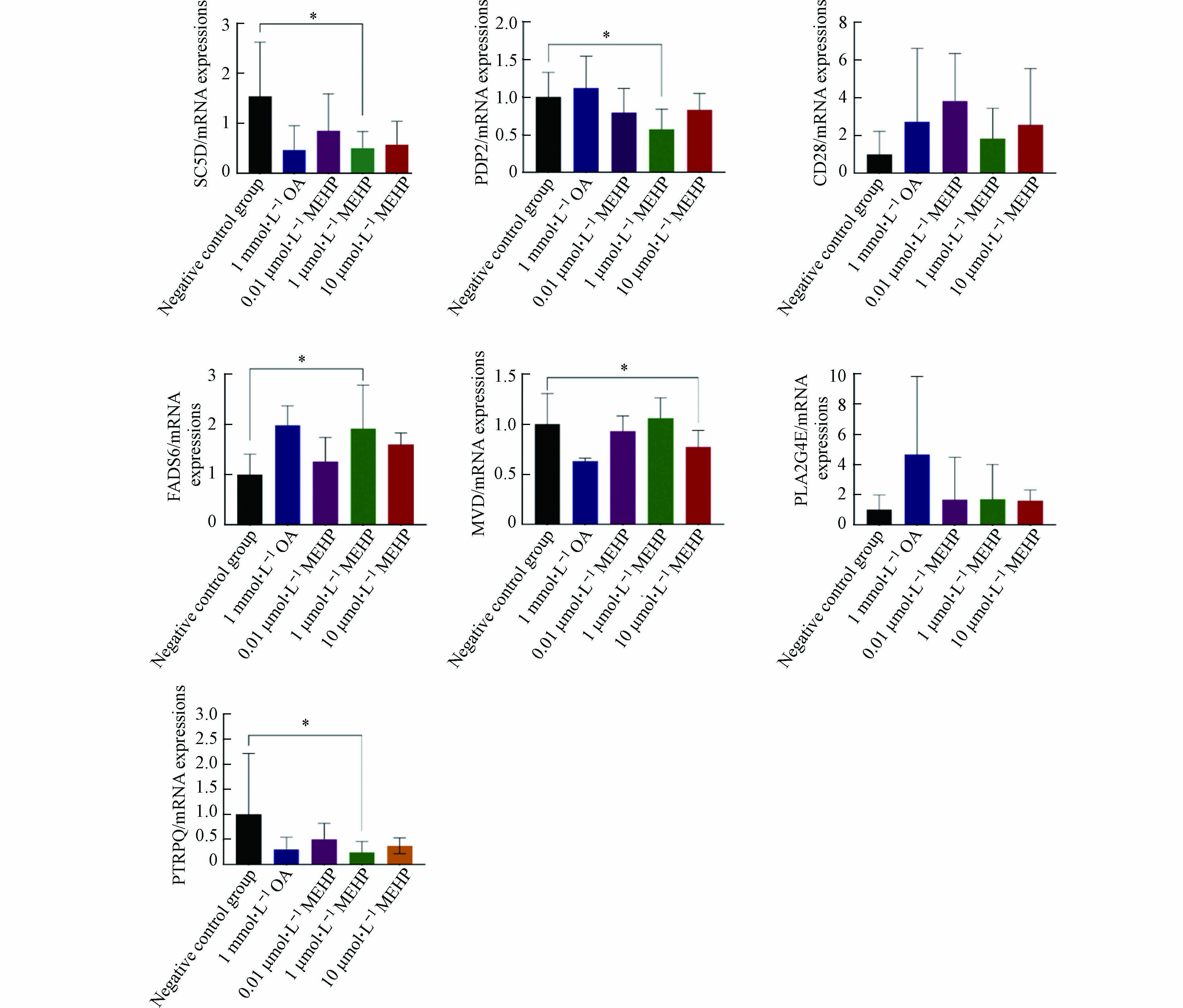
图 4 不同浓度MEHP染毒对 HepG2 细胞相关基因mRNA表达水平影响
Figure 4. Effects of different concentrations of MEHP on mRNA expressions of relative genes in HepG2 cells
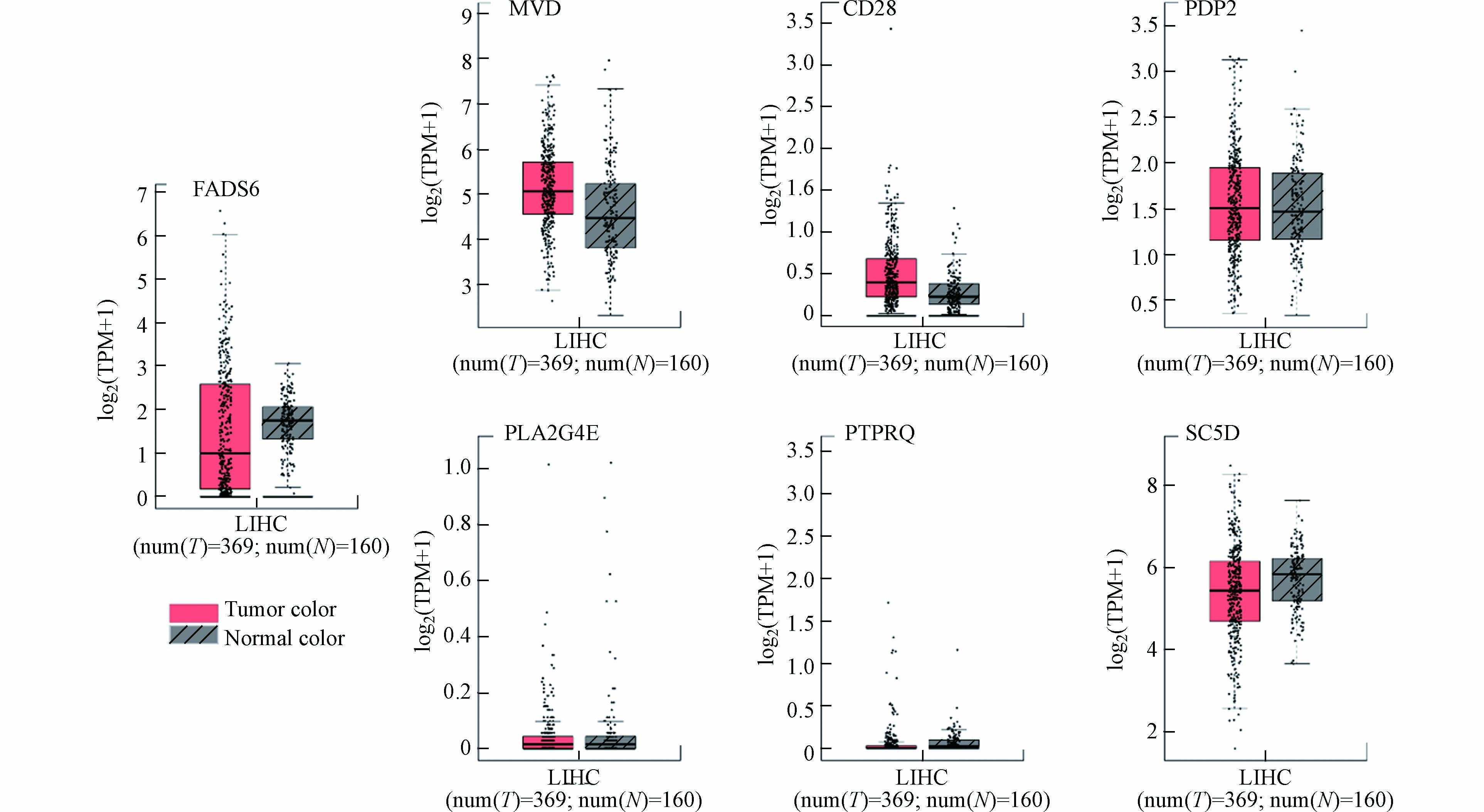
图 5 关键基因在肝癌中的表达水平
Figure 5. Relative expression comparison of key genes in liver cancer samples in GEPIA database
表 1 实时定量PCR引物序列
Table 1. Primer sequences for real-time quantitative PCR
基因
Gene引物
PrimerPDP2 S——GAAGATGAGGTGACAAGGAA F——GCCAGCACAAG MVD GAACTTA PLA2G4E S——TTCTGTCCTATGGCTCCTT F——GTTCTTCACTCGGCTCTG FADS6 S——CCTCAACCGCTATGTCTAC F——CGATGTGCTGGAAGATGT PTPRQ S——ATGTCTATATTGCGGCTGAA F——TTCTTACTTGCGTGGATTCT CD28 S——GCTCTTGGCTCTCAACTTA F——CCTGCTCCTCTTACTCCT SC5D S——CTTGCTGGAGATAAGAGGTT F——TATGGTGGTCTGTATGATGAG MVD S——CAAGGACTTCACCGAGGA F——GTAGGCTAGGCAGGCATA  下载: 导出CSV
下载: 导出CSV
表 2 差异基因的表达情况
Table 2. The expression of differentially expressed genes
Up Down symbol lgFC P.Value symbol lgFC P.Value DPPA3 0.743871 0.000032637 TTTY14 −0.75345 0.000683968 MYLIP 0.685596 0.000320629 XLOC_008352 −0.691 0.001413602 XLOC_000888 0.664559 0.000498704 LOC100129112 −0.83639 0.001799658 LOC100127994 0.609364 0.00056601 SC5D −0.92097 0.002090916 SYT17 0.650856 0.000986627 XLOC_004178 −0.72657 0.002192512 XLOC_008003 0.727595 0.00115933 C14orf119 −0.78947 0.003595082 LOC100506487 0.589801 0.001543009 XLOC_l2_012082 −0.61338 0.004605101 FADS6 0.784036 0.002373877 XLOC_012908 −0.92876 0.006283058 AQP10 0.603687 0.00355145 GABRG1 −1.01945 0.007994 XLOC_001387 0.675839 0.003916862 NUMB −0.57939 0.008775398 KCNE4 0.831973 0.003949384 FLJ21408 −0.61905 0.009899712 XLOC_005215 0.65671 0.004608305 LOC100505657 −0.89616 0.010826125 OR5L1 0.922639 0.004609802 ZNF554 −0.60913 0.011699612 C14orf180 0.649561 0.005207664 XLOC_012871 −0.78553 0.01385829 RHOXF1 0.622636 0.007822227 XLOC_007131 −0.7273 0.014006601 PTPRQ 0.739332 0.008548009 XLOC_l2_014785 −0.66169 0.01855114 LOC286382 0.586815 0.008726912 PRO0611 −0.78996 0.018748849 ACOD1 0.61305 0.008957375 CREG2 −0.70749 0.018758137 LOC100130744 0.85844 0.009748146 XLOC_l2_004857 −0.66225 0.023632428 ZAP70 0.625018 0.010183849 PDP2 −0.80976 0.023933353 PLA2G4E 1.427762 0.011394311 XLOC_l2_011207 −0.57923 0.026335268 LOC652586 0.614123 0.011818286 XLOC_007761 −0.58384 0.027416794 XLOC_006019 0.666199 0.012002598 GNL3LP1 −0.6286 0.027632132 SLAMF7 0.896594 0.013447959 XLOC_l2_013646 −0.89314 0.028992205 XLOC_005433 0.588529 0.014072632 MVD −0.63712 0.031037423 KRTAP13-2 0.681023 0.014111061 LOC728065 −0.58089 0.031598939 XLOC_008237 0.804098 0.01445258 XLOC_l2_011011 −0.60147 0.033573436 XLOC_008244 0.677154 0.01652238 XLOC_001687 −1.03697 0.037696163 LOC100505966 0.836326 0.01818776 XLOC_004804 −0.68571 0.039579631 XLOC_001422 2.454291 0.018227482 LOC100507110 −0.60107 0.041523521 CSN2 1.0231 0.020348553 CACNG8 −0.61314 0.041704333 XLOC_003782 0.639271 0.021116476 SNORD70 −0.92894 0.044259091 XLOC_005572 0.585382 0.02204518 LOC100128126 −1.05439 0.044856832 XLOC_003349 1.233979 0.022738339 NPPA-AS1 −0.59523 0.048087643 XLOC_l2_00770 0.610547 0.023106187 XLOC_001550 −0.64752 0.04894801 SNORD115-48 0.719119 0.024508996 INE1 −1.017 0.0496582 SNORD115-4 0.688309 0.026211298 XLOC_001755 0.605516 0.028061446 ANK2 0.906401 0.029169596 CD28 0.589859 0.029610282 LOC100506563 1.019834 0.029837404 ANXA10 0.626939 0.030222491 CYBB 1.151916 0.03336549 XLOC_012064 1.019834 0.0345969 XLOC_013649 0.624561 0.035226312 XLOC_010183 1.380268 0.035959747 XLOC_006983 0.862151 0.040708371 XLOC_004274 0.720596 0.040795527 XLOC_008690 0.686422 0.041491936 XLOC_007888 0.852148 0.04152697 XLOC_l2_011145 0.704819 0.042378972 SLC24A4 0.757531 0.043335006 XLOC_013458 0.71734 0.043999913 TREML1 0.800135 0.047577606 TRIM49 0.796312 0.048328175 ANKDD1B 0.683738 0.049447534 DUSP21 0.604901 0.049510457
下载: 导出CSV
表 3 DAVID分析中前5个通路(BP)与基因
Table 3. Top 5 GO terms (BP) of the genes with the DAVID analysis
GO Term
GO 条目Count
数量Genes
基因Fold Enrichment
富集倍数P Value
P值GO:0044255
cellular lipid metabolic process7 PDP2, PLA2G4E, FADS6,
PTPRQ, CD28, SC5D, MVD3.85628326776209 0.0068 GO:0045087
innate immune response5 TREML1, ZAP70, CYBB,
ACOD1, SLAMF73.042438312 0.0494 GO:0055085
transmembrane transport7 SLC24A4, CACNG8, KCNE4, AQP10,
CYBB, ANK2, GABRG13.024137931 0.0204 GO:0006629
lipid metabolic process7 PDP2, PLA2G4E, FADS6, PTPRQ,
CD28, SC5D, MVD5.660947851 0.0210 GO:0034762
regulation of transmembrane transport4 CACNG8, KCNE4, CYBB, ANK2 3.43652037617554 0.0300
下载: 导出CSV
-
[1] KAMRIN M A. Phthalate risks, phthalate regulation, and public health: A review [J]. Journal of Toxicology and Environmental Health, Part B, 2009, 12(2): 157-174. doi: 10.1080/10937400902729226 [2] CIRILLO T, FASANO E, ESPOSITO F, et al. Di(2-ethylhexyl)phthalate (DEHP) and di-n-butylphthalate (DBP) exposure through diet in hospital patients [J]. Food and Chemical Toxicology, 2013, 51: 434-438. doi: 10.1016/j.fct.2012.10.015 [3] COLACINO J A, HARRIS T R, SCHECTER A. Dietary intake is associated with phthalate body burden in a nationally representative sample [J]. Environmental Health Perspectives, 2010, 118(7): 998-1003. doi: 10.1289/ehp.0901712 [4] 高海涛, 李瑞仙, 邸倩南, 等. 我国人群邻苯二甲酸酯类的暴露水平及风险 [J]. 癌变·畸变·突变, 2017, 29(6): 471-475. GAO H T, LI R X, DI Q N, et al. Exposure level and risk of phthalate esters in Chinese population [J]. Carcinogenesis, Teratogenesis & Mutagenesis, 2017, 29(6): 471-475(in Chinese).
[5] ROWDHWAL S S S, CHEN J X. Toxic effects of di-2-ethylhexyl phthalate: An overview [J]. BioMed Research International, 2018, 2018: 1750368. [6] BHATTACHARYA N, DUFOUR J M, VO M N, et al. Differential effects of phthalates on the testis and the liver [J]. Biology of Reproduction, 2005, 72(3): 745-754. doi: 10.1095/biolreprod.104.031583 [7] 聂燕敏, 齐丽娟, 高珊, 等. 邻苯二甲酸酯对健康影响的危害评估 [J]. 毒理学杂志, 2021, 35(5): 361-366. doi: 10.16421/j.cnki.1002-3127.2021.05.001 NIE Y M, QI L J, GAO S, et al. Hazard assessment of the effects of Di-(2-ethylhexyl) phthalate (DEHP) on health [J]. Journal of Toxicology, 2021, 35(5): 361-366(in Chinese). doi: 10.16421/j.cnki.1002-3127.2021.05.001
[8] 贺真. 邻苯二甲酸单乙基己基酯对HepG2细胞甘油三酯合成的影响[M]. 太原: 山西医科大学, 2019. HE Z. Effects of Mono (2-ethylhexyl) Phthalate on the Synthesis of Triglyceride in HepG2 Cells[M]. Taiyuan: Shanxi Medical University, 2019(in Chinese).
[9] YANG G T, ZHANG W J, QIN Q Z, et al. Mono(2-ethylhexyl) phthalate induces apoptosis in p53-silenced L02 cells via activation of both mitochondrial and death receptor pathways [J]. Environmental Toxicology, 2015, 30(10): 1178-1191. doi: 10.1002/tox.21990 [10] SCHWARTZ M W, SEELEY R J, ZELTSER L M, et al. Obesity pathogenesis: An endocrine society scientific statement [J]. Endocrine Reviews, 2017, 38(4): 267-296. doi: 10.1210/er.2017-00111 [11] THOMAS J A, NORTHUP S J. Toxicity and metabolism of monoethylhexyl phthalate and diethylhexyl phthalate: A survey of recent literature [J]. Journal of Toxicology and Environmental Health, 1982, 9(1): 141-152. doi: 10.1080/15287398209530149 [12] KWACK S J, HAN E Y, PARK J S, et al. Comparison of the short term toxicity of phthalate diesters and monoesters in sprague-dawley male rats [J]. Toxicological Research, 2010, 26(1): 75-82. doi: 10.5487/TR.2010.26.1.075 [13] JIA Y Y, LIU T, ZHOU L T, et al. Effects of di-(2-ethylhexyl) phthalate on lipid metabolism by the JAK/STAT pathway in rats [J]. International Journal of Environmental Research and Public Health, 2016, 13(11): 1085. doi: 10.3390/ijerph13111085 [14] SEO K W, KIM K B, KIM Y J, et al. Comparison of oxidative stress and changes of xenobiotic metabolizing enzymes induced by phthalates in rats [J]. Food and Chemical Toxicology, 2004, 42(1): 107-114. doi: 10.1016/j.fct.2003.08.010 [15] CUI J, CHEN H Q, TANG X, et al. Δ6 fatty acid desaturases in polyunsaturated fatty acid biosynthesis: Insights into the evolution, function with substrate specificities and biotechnological use [J]. Applied Microbiology and Biotechnology, 2020, 104(23): 9947-9963. doi: 10.1007/s00253-020-10958-5 [16] TOSI F, SARTORI F, GUARINI P, et al. Delta-5 and delta-6 desaturases: Crucial enzymes in polyunsaturated fatty acid-related pathways with pleiotropic influences in health and disease [J]. Advances in Experimental Medicine and Biology, 2014, 824: 61-81. [17] MONTELL E, TURINI M, MAROTTA M, et al. DAG accumulation from saturated fatty acids desensitizes insulin stimulation of glucose uptake in muscle cells [J]. American Journal of Physiology. Endocrinology and Metabolism, 2001, 280(2): E229-E237. doi: 10.1152/ajpendo.2001.280.2.E229 [18] SONG T X, WANG P L, LI C Y, et al. Salidroside simultaneously reduces de novo lipogenesis and cholesterol biosynthesis to attenuate atherosclerosis in mice [J]. Biomedicine & Pharmacotherapy, 2021, 134: 111137. [19] JUMP D B, BOTOLIN D, WANG Y, et al. Fatty acid regulation of hepatic gene transcription [J]. The Journal of Nutrition, 2005, 135(11): 2503-2506. doi: 10.1093/jn/135.11.2503 [20] YAPLITO-LEE J, PAI G, HARDIKAR W, et al. Successful treatment of lathosterolosis: A rare defect in cholesterol biosynthesis—A case report and review of literature [J]. JIMD Reports, 2020, 56(1): 14-19. doi: 10.1002/jmd2.12158 [21] HO A C C, FUNG C W, SIU T S, et al. Lathosterolosis: A disorder of cholesterol biosynthesis resembling smith-lemli-opitz syndrome [J]. JIMD Reports, 2014, 12: 129-134. [22] COLAK Y, SENATES E, OZTURK O, et al. Association of serum lipoprotein-associated phospholipase A2 level with nonalcoholic fatty liver disease [J]. Metabolic Syndrome and Related Disorders, 2012, 10(2): 103-109. doi: 10.1089/met.2011.0111 [23] 韩晓蕾. Lp-PLA2对肝脏细胞脂代谢的影响[D]. 长春: 吉林大学, 2016. HAN X L. Effects of lp-PLA2 on lipid metabolism of hepatocyte[D]. Changchun: Jilin University, 2016(in Chinese).
[24] TSIKAS D, SANDMANN J, ROSSA S, et al. Investigations of S-transnitrosylation reactions between low- and high-molecular-WeightS-nitroso compounds and their thiols by high-performance liquid chromatography and gas chromatography-mass spectrometry [J]. Analytical Biochemistry, 1999, 270(2): 231-241. doi: 10.1006/abio.1999.4084 [25] 代雪枫, 王洪飞. 非酒精性脂肪肝病与肝癌 [J]. 实用全科医学, 2007, 5(7): 647-649. DAI X F, WANG H F. Nonalcoholic fatty liver disease and liver cancer [J]. Applied Journal of General Practice, 2007, 5(7): 647-649(in Chinese).
[26] LIOU I, KOWDLEY K V. Natural history of nonalcoholic steatohepatitis[J]. Journal of Clinical Gastroenterology, 2006, 40 Suppl (1): S11-S16. [27] ERTLE J, DECHÊNE A, SOWA J P, et al. Non-alcoholic fatty liver disease progresses to hepatocellular carcinoma in the absence of apparent cirrhosis [J]. International Journal of Cancer, 2011, 128(10): 2436-2443. doi: 10.1002/ijc.25797 -
 点击查看大图
点击查看大图
计量
- 文章访问数: 5259
- HTML全文浏览数: 5259
- PDF下载数: 115
- 施引文献: 0
